
The problems of species classification of Haworthia should now be well known to all enthusiasts of this interesting genus. I have proposed and maintained, with cosmetic changes, a nomenclatural system for it since 1975. It is a system with which I have managed an extensive collection and herbarium record, and I know it works within the limitations imposed by the evident fractal nature of “species” and their variability. In this paper I would like to expose these limitations with respect to the concept of two species viz. Haworthia mirabilis, and Haworthia maraisii, where there may be only one. (In the original hardcopy publication of this article, the illustrations are all captioned H. maraisii when it would have been sensible to have used H. mirabilis).
In most discussions concerning the classification of Haworthia, participants have suggested that there are too many species and that some of them should be “lumped”. On the other hand, there have been several writers who, as prospective taxonomists and experts on the group, continue to expand the range of entities at the formal rank of species and varieties despite all the evidence and indications that this could be an endless path leading nowhere. My own inclinations have been to minimize the number of species and to use varietal rank in two ways as a communication medium. One is that I have tended to reject varieties of older authors which I did not think had a strong enough geographical base, but also recognizing that there is nevertheless some information inherent in such names. Often I felt these varieties simply expressed the variability in a way that was insignificant with respect to the species as a whole. Two, is that I have described some new varieties to provide names for morphological variation that I consider is new and previously unrecognized, accepting that these names should perhaps not be immortalized either. Thus my proviso has been that these new varietal names should not be taken too seriously. As far as the number of species is concerned, I know full well that there could be fewer species. I get caught up in the problem of identifying a “species” in a strict botanical definition of the word, as opposed to the need for “names” as a way of simple communication about the plants in the amateur fraternity. Because of the problem of similarity and continuity, the elimination of names becomes similar to that of falling dominoes and the question then arises of “where does it end?” My classification should not be confused with a system of names intended for horticulture or for trade. But neither do I think such a system should adulterate a formal botanical one.
Therefore, underpinning this presentation is a definition of a species as dynamic systems of living organisms morphologically, genetically and even behaviorally, continuous in space and time.
What is an Haworthia species? I do not accept that there are different kinds of species nor do I accept the term “biological species”. A species is a biological construct intended for life form and the classification of Haworthia has been presumably intended to identify those individual species that constitute a genus set. It seems evident that the commonest element of any species concept, expressed or unexpressed, is that of reproductive isolation i.e. members of different species cannot interbreed. In Haworthia this evidently does not apply because there is compatibility between individual plants of even different sub-genera, what to say of intergeneric hybridization. If there are breeding barriers they are the obvious isolationary ones that are regarded by zoologists as prelude to speciation i.e. geographic isolation, behavioural (different flowering time) and genetic (polyploidy) factors. Genetic or other breeding complexities may, in my opinion, be characteristics within such sets as much as they help define them. In that respect I do consider that geographical (and geological) breeding and survival barriers are in evidence and are significant. My perception of the species is then of sets of individuals and populations which suggest morphological and genetic continuity in time and space
The problem of continuity is nothing more than the assessment of similarity and difference. This point needs examination, as it is basic to the question of characterization and classification. The species is the basic biological unit used to identify life forms. In the higher life forms (animals) it seems to be fairly straightforward and what could be problems are easily resolved e.g. the lion species is essentially African, but there is an Asian representative. This is similarly the situation of the elephant species. In one case are they classified as different species, in the other, two. Zoology recognizes and places strong emphasis on the capacity of animal life forms to interbreed and produce fertile offspring. In the lower life forms there are problems because interbreeding capacity does not seem to relate so closely to the life forms and fertile hybrids are produced from improbable parentage. However, it is unlikely that this is the criterion that is actually used to identify “species”. In my opinion reproductive isolation in a species definition has been a by-product of evolutionary philosophy and that botanists have simply assumed that a species definition for animals also holds for plants.
Continuity is an aspect that I do consider in relation to occurrence with other species and in relation to geography. My observations have been that plants within populations, and thus the populations themselves, show trends. These trends are referred to in botany and zoology as clines and it does happen that the tails or two ends of such a cline may meet and thus produce a situation where a classification decision for one geographic area, may not be correct for another. Then there is the problem of skeletal soils and habitat difference and the effect this has on plant appearances – either generated simply by different soils and/or due to differential selection on the basis of genetic characters. It seems to me simply obvious that sometimes the quite apparent differences between two or more groups of plants is due to local differences in geology and has no general application to the same plants over a greater geographical range. Perhaps it is worth admitting Darwin’s observation to be correct: any local truth or integrity of species is greatly reduced over large areas, and perhaps extend the idea that it is also very greatly reduced by the amount of material examined.
Ideally similarity or difference should be measurable and this is manifested in numerical taxonomy whereby measurements of every conceivable feature are accumulated and processed statistically. It is not possible for me to generate such statistical data in any conventional way and I have to rely on perception derived from learning and experience of what constitutes “difference”. Collecting permits only allow three specimens and it would in any case be ridiculous to suggest that more plants than this should be gathered simply for statistical demands. This learning of mine has been done not only within the limits of Haworthia, but within animal (largely insect) and many other plant genera.
How can a Haworthia species be circumscribed? Description plays a critical role in the examination of similarity. The irony is that if one examines all the descriptions relating to the taxa (species and varieties) that I will cover in this article, it is patently obvious that descriptive detail is practically worthless. Detail barely enables one to describe a single specimen with confidence that it could be identified among others on the basis of its description, and this is even less likely if the growing conditions of that one plant are changed. It has been difficult even within the scope of this one study to photograph plants that are strictly comparable in terms of growing condition. We are also faced with the fact that individual plants within populations are highly variable and that the description of a single plant is far from the description of a population. Furthermore, the description of populations is also compounded by numbers and the difficulty of arriving at a norm by which it would be reasonably possible to identify individuals of any one population of few or many.
Objective? Recognizing the above limitations, this article is intended to examine whether or not H. mirabilis and H. maraisii, as I presented them in my revision (Haworthia Revisited, 1999), can be shown to be separable sets of individuals and populations. I wish to illustrate H. mirabilis and the interplay of this species with H. maraisii as I have experienced it in and around the Riviersonderend Mountains. I conclude that they are but one species and within that decision is nested the problem of what then of related species and where does it end? This is because the whole story can barely be told in one piece as both elements have extended distributions where they unfold other variance and interact with other “species”. Thus other names have also been used in this context and there is a plethora of such names associated with the problem of characterization and classification that need to be understood.
History and background information:
Although the name H. mirabilis has a long taxonomic history dating from the original description by Haworth in 1804, the name was curiously not associated with field plants until I did so in 1973 (Haworthia Handbook). In the interim several other species (and varieties) were described which originated out of what is obviously a single complex in the greater Bredasdorp/Caledon area. These species included names such as mundula, badia, triebneriana, eleven varieties of triebneriana, and emelyae var. beukmannii. Typification of the name is quite easy if the exact original locality is known. But it is difficult for the nomenclatural type to be typical of any taxon for the reason that in Haworthia, the plants grow in scattered and isolated populations. There is variability and difference of plants both within and between these populations. Often one can be completely certain from which population a plant may have come, and in other cases it is not even certain from which set of populations a plant may have come. Thus it was that the name mirabilis had not been associated with any natural source until I did so, and then I felt constrained to conclude that the population from which it most probably had come was that from which G.G. Smith had drawn his H. mundula. This particular population is very characteristic and individuals can be identified with a very high degree of certainty. The consequence is that we have a situation where the species name is based on plants that are not truly representative of the others in the set. Many of the other variants were described under the name H. triebneriana. Some of these can be identified with a high degree of certainty and others not. The name triebneriana seems to be associated with a population at Stormsvlei (it is certain that Von Poellnitz wrongly used the name Strydomsvlei) and there are two other varieties that also had this place as source – they are H. triebneriana vars depauperata and pulchra. This suggests that the var triebneriana cannot be identified with certainty. The variety beukmannii was attached to the species H. emelyae, and this also indicates how indeterminate the species were in the absence of reasonable definition and capability.
A similar situation exists with regard to the name maraisii which was a species originally also collected at Stormsvlei. We only know this because it was originally reported in the description as having come from “near Swellendam”, and it was G.J. Payne who showed me the place at Stormsvlei. It was I who extended the use of the name to a wider range of populations, as I will discuss in this paper. I have said elsewhere (Haworthiad 17:83, 2003) that there was an unfortunate decision by W.T. Stearn to typify the name herbacea on the basis of an old illustration (Boerhaave, 1720) that was better associated with the name atrovirens. I unfortunately applied this name herbacea according to the descriptive content of the name (yellow-green in reference to spines) and used it for a species in the Worcester/Robertson area. The name atrovirens, the meaning of this name (dark green referring to leaves), and the accompanying original illustration could perhaps have been better applied to the species which von Poellnitz named H. maraisii and also to H. schuldtiana. The species name H. maraisii thus originates at Stormsvlei and I have used the name for a series of smallish dark-green plants occurring mostly north of the Riviersonderend Mountains in the Robertson area. I did this because I saw a similarity of the plants at Stormsvlei to these Robertson Karoo elements. Von Poellnitz did not make this connection and applied the name H. schuldtiana (and of course several varieties) to most of the collections from north of the Riviersonderend just as he applied the name H. triebneriana (plus several varieties) to most of the collections south of the mountains. The problem of certain identification of names is again prominent and the schuldtiana varieties minor and subtuberculata are from the same locality north of McGregor. Again H. schuldtiana was described simply as “Robertson”. The collector was G.J. Payne and he pointed out to me the true origin west of McGregor.
There is a problem in the way the name H. maraisii has been typified and then applied by me. When collected it was reported by W.R.B. Marais to have come from “weathered slate crevices on hills facing the Karoo near Swellendam”. J.R. Brown reported having received a plant from W.E Armstrong from Stormsvlei but this is perhaps from north of the Pass. G.J. Payne showed me the locality where Marais was purported to have actually collected the plant and I assumed Payne was correct as the plants there agreed closely with the illustration in the Berlin herbarium taken to be the type. This locality is south of the Stormsvlei Pass, and also south of the river, in shale outcrop among what is now a complex of buildings. The site is barely intact but while there were plants there in 1970, I was not been able to locate them on succeeding visits, although I have subsequently done so. In my Handbooks I cited a plant from north of McGregor as “a representative specimen” but indicated that I accepted the illustration in the Berlin herbarium as the type. Breuer, whose typifications I regard as untimely and unfortunate, makes a confusing statement in his first book “World of Haworthias Vol.1”. Here he cites my selection of the Berlin photograph as lectotypification of the name H. maraisii but cites the locality of my representative specimen from McGregor as the original locality of that species. He also maintained that as the Berlin specimen “is dated from 1937”, it is “therefore no original material”. In his “World of Haworthias Vol.2” he has changed his mind about the date of the Berlin illustration and notes it as “undated” and apparently accepts the fact that this is the valid type. However, he uses a plant from his own collection IB5926 from Swellendam to also illustrate the species. The plant IB5926 looks very similar to clones in my own collection MBB6644 (and/or 6860, populations 10 and 11 in the text below) that comes from close to Swellendam and some way from Stormsvlei. The problem for me is that I associate the population at Swellendam (there are two similar populations) with a matrix of populations and individuals (Set A to follow) that I cannot classify with any degree of certainty. Thus Breuer may have unintentionally and inadvertently generated the end-point that I want to achieve with this paper, indicating that maraisii and mirabilis are the same thing by virtue of substituting a mirabilis-like plant for a maraisii-like one. It will transpire that my decision to use the name H. maraisii also for the complex north of the Riviersonderend Mountains viz. H. schuldtiana, is an identical misstep. This is because I revisited Stormsvlei while finalizing this manuscript and fortuitously found plants again very close to the original spot. While these plants do indeed appear to be very different to H. mirabilis as it occurs within the Stormsvlei Pass, it is now quite evident on the basis of experience and re-examination that the difference is due to the geological substrate on which the populations occur. The connection with populations north of the Riviersonderend Mountains, specifically to the one at McGregor, is still true but it means that I have used a name based on an ecotype of mirabilis viz. maraisii, when it would have been more appropriate to have used the name schuldtiana.
Considerations:-
a. The classification of von Poellnitz (1940):-
The basic difference between the two species as recognized by von Poellnitz can be evaluated from a key which he produced and which was published in Fedde’s Rep.43:102 (1938). The main feature or character of his section Retusae in which he placed the two species was “leaves more or less erect, with minute teeth on the margin, more rarely smooth, at the top bent back sharply and resulting in an end-area which is somewhat translucent, with a few or more green stripes, smooth, tubercled or barely with minute teeth”. In this key von Poellnitz separated H. schuldtiana (with his H. paradoxa) on the basis of the end surfaces of the leaves with small teeth and tubercles. Species with the end-surfaces tubercled only or smooth included H. mirabilis together with H. triebneriana and H. willowmorensis, which I regard as synonymous. Also included here were H. maraisii, H. sublimpidula and H. whitesloaneana. I have explained that the former can be regarded as an ecotype of H. mirabilis. This could confound my use of the name as synonymous with H. schuldtiana where the latter two belong. He also includes in his key H. paradoxa and H. magnifica but these are two elements from east of the Breede River and it will become apparent that I exclude them from this discussion on the basis of geographical consideration only. In my Revision (1999) I have actually regarded H. paradoxa as a variety of H. mirabilis while H. magnifica is upheld as a distinct species despite the obvious affinity with the two species being discussed in this work. Although he also described H. badia in 1938, von Poellnitz did not include this species in his key to the Retusae where it unquestionably belonged.
In total Von Poellnitz’ key separates 23 “species”. He published a key to nine varieties of H. triebneriana (two more were added in 1940) in Feddes’Rep. 47:1 (1939). In the same journal 49:23 (1940) he published a key to 10 varieties of H. schuldtiana that included the former H. whitesloaneana and also what I upheld as H. maculata. My observations are that it is not possible to use the characters von Poellnitz used to separate the varieties as geographic entities by virtue of the inherent variability between plants even within single population.
b. The classification of Bayer (1999):-
I considered that H. mirabilis and H. maraisii constituted sets on the basis of geographical distribution. Thus in the southwest the plants are larger or more substantial, with erect leaves often spotted on the back or under surfaces, and with fairly conspicuous marginal spines i.e. H. mirabilis. In the northwest the plants are smaller and darker green, the leaf tips tend to retuse and the marginal spines are reduced i.e. H. maraisii. I did recognize a color difference in that H. mirabilis was generally reddish in color with translucence in the leaf end-area, as opposed to the dark-green non-translucence of H. maraisii. In essence, Von Poellnitz’ key of 1940 suggests that the two elements I recognize were distinguishable on the basis of “end surfaces with small teeth and tubercles” in his H. schuldtiana (included in my maraisii) and “end surface with tubercles only or smooth” in H. mirabilis AND in H. maraisii (excluded from my broader use of the name). This is only partly true in that while such a tendency might be there, it is not an infallible distinction.
My opinions were simply generalisations derived from a body of populations, one south and one north of the Riviersonderend Mountains. The interplay of these two species, H. mirabilis and H. maraisii, is limited by the fact that the occurrence of mirabilis is at the extreme southwest of the distribution range of the subgenus Haworthia. At its eastern limits there may be some interaction with H. rossouwii in the south, and perhaps even with H. mutica. But otherwise the interaction is with H. maraisii (as I apply the name) and the flow of that species westward and southward with H. heidelbergensis and H. floribunda. It is not my intention to examine that whole complex issue here. What I will show is that across the Riviersonderend Mountains, H. mirabilis becomes inextricably interconnected with H. maraisii and perhaps even with H. maculata. Not only that, it will appear that at Stormsvlei, where H. maraisii, and H. mirabilis(triebneriana) vars. pulchra and depauperata originated, Haworthia maraisii and H. mirabilis can be assumed to be ecotypes of the same one species because of the proximity of the habitats and the synchronization of flowering time and seed set. The only reasonable inference seems to be that the plant differences, which are essentially minimal, are due to the dramatic differences in the skeletal soils and exposed rock they are associated with.
The overriding complexity is that the core of the H. maraisii/mirabilis relationship is expressed in a small series of populations that extends from Heidelberg in the east to Riviersonderend in the west. This includes the original site for H. maraisii at Stormsvlei. It should be borne in mind that I also initially regarded H. maraisii and H. magnifica as one and the same. While I uphold them as separate species in my revision I only do so to facilitate recognition of variants in such an east/west split. I cannot explain the situation presented by populations of plants down the Duiwenhoks river which have to some degree been discussed elsewhere (see Chapter 6). The interplay of H. maraisii with other taxa is a little more complex and I will deal with the issue by examining populations as sets, either in this presentation or separately (I have written a discussion on the H. heidelbergensis/H. magnifica interaction east of Heidelberg also Chapter 6, and also a discussion of the variation and distribution of H. floribunda¸ Chapter 1).
c. Skeletal soils, geology and ecotypification:-
I have shown (Chapter 8, Update 2) that in the E. Cape geology plays a very significant role in the way the species vary and relate to one another. My original observations that this was true were made in the W Cape and Little Karoo and I have no doubt that this is very true for the species now being considered. It is probably true for the Cape winter rainfall biomes (I use the plural only because professional botanical wisdom maintains that there are more than one) that the floras are so species rich because of skeletal soils, geological and topographical diversity and strong rainfall differences (with reliable, consistent and effective winter rain). My study area in this case is dominated by rock formations of the Cape Supergroup although in the north the Karoo Supergroup makes its appearance. The Cape Supergroup comprised three groups viz. Table Mountain Sandstone, Bokkebeld Shales and Witteberg Sandstones and these comprise again about 27 formations. These rocks are invariably directly exposed and soils are often very skeletal. Rock is almost a prerequisite for the presence of Haworthia and it is quite evident to even the most casual observer that geology and geomorphology must in some way impact on the evolutionary processes and pressures within the genus whatever they are. But while geology is no doubt significant, I will only cite it here and there as a rough pointer. This is because although the rocks in the Cape supergroup vary from pure sandstone in the Table Mountain group to mudstones in the Bokkeveld group and again sandstones in the Witteberg group, there are many variants which are obscured at the general map scale of 1:50 000. There is apparently also no general rule that a particular vegetative form will repeat itself on identical rock formation or soil.
Further east in the Riversdale and Heidelberg areas, ferricrete inselbergs are significant Haworthia habitats. As can be seen from my map, the area between Swellendam and Bredasdorp where such inselbergs are abundant, there are almost no records for Haworthia. This could be due to poor exploration. On the other hand H. mutica does occur there and the void may be partly explained by virtue of the occurrences of a different species. To some extent the occurrence of an element which I refer to as H. heidelbergensis is used to fill a similar void in the south and southeast. On the accompanying distribution map, other species occurrences are thus also indicated.
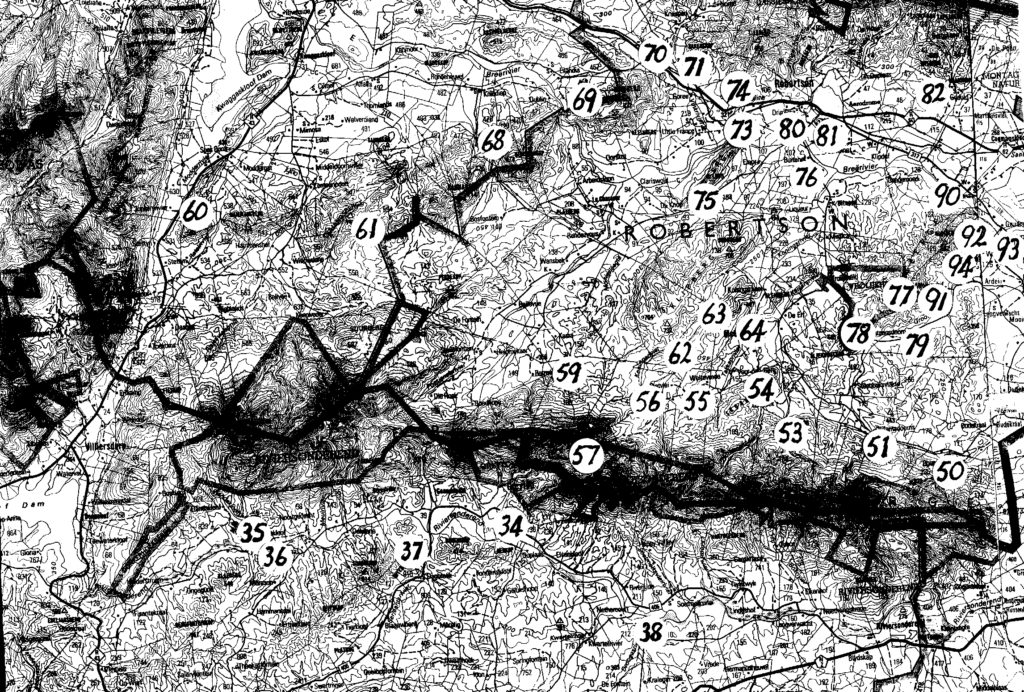
Map 1. Worcester to Robertson 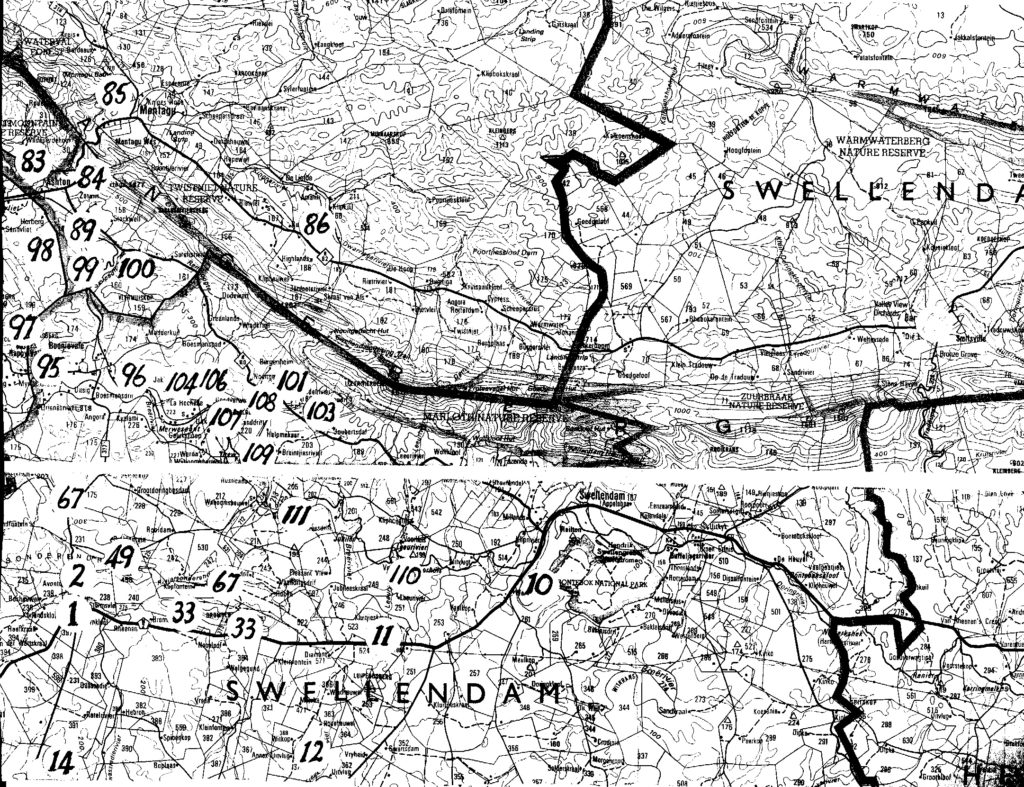
Map 2. Ashton to Swellendam. 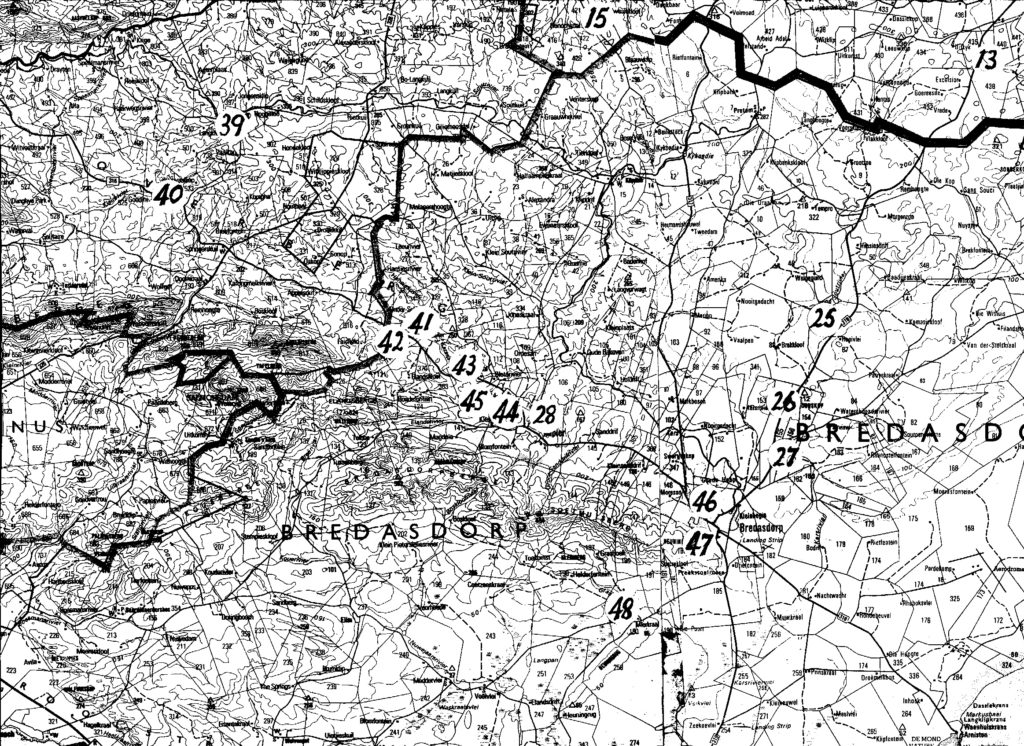
Map 3. Bredasdorp 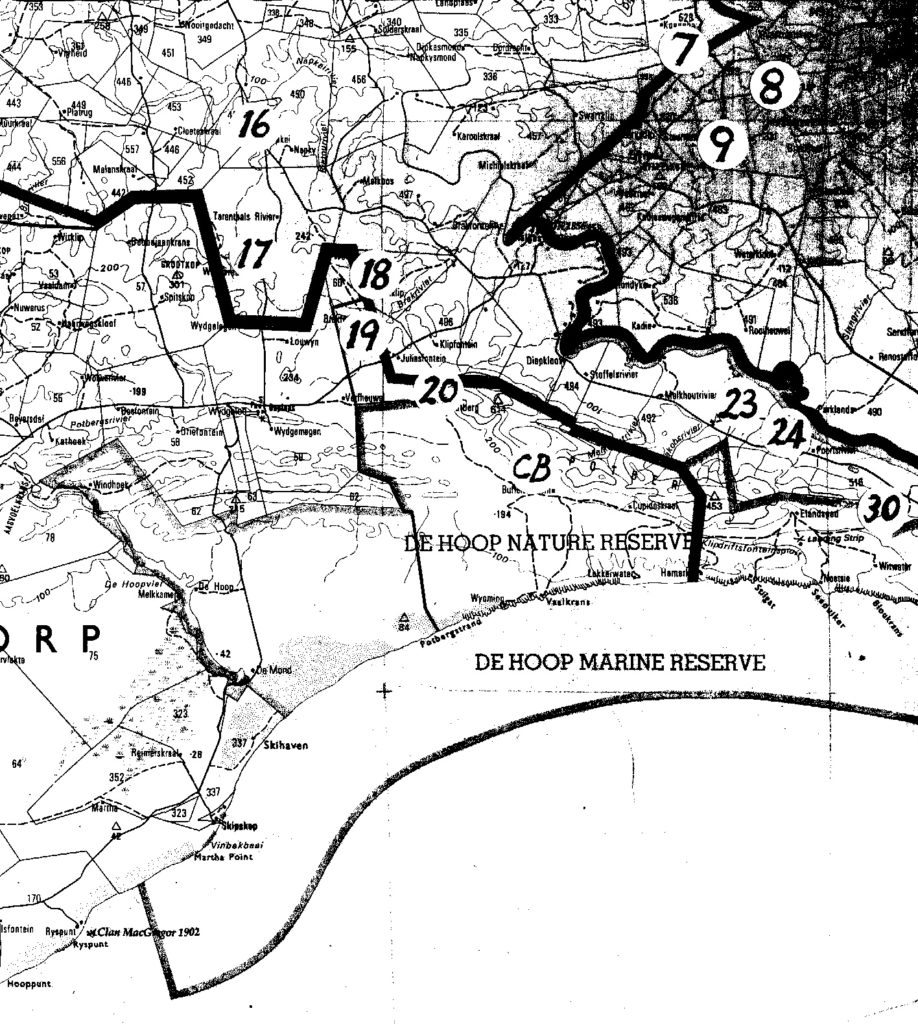
Map 4. E Bredasdorp.
The population sets examined:-
The entire problem is summarized in the listing of six sets of populations (A…F). The location of these is presented on the accompanying map. Because of scale, a mapped site may indicate 2 populations and the sequential population number then omitted. The plants are illustrated and numbered in the same sequence that the populations are listed:-
A. Haworthia maraisii south and east of Riviersonderend Mountains:-These constitute a geographic set which occupies the southeastern range of the study area. A principle boundary is the Breede River but I have included six populations from further east of which four are off the map. Those four populations will show the integration of the sets considered here, with sets east of Swellendam. The plants are generally dark-green with reduced spination. In this area H. maraisii becomes inextricably linked with what I regard as H. heidelbergensis. That in turn has a complex interplay with H. floribunda in the southeast, with H. turgida in the east and then with H. magnifica still further east. There is a complication too in terms of a few populations along the south bank of the lower Breede River near Malgas and Infanta that is possibly an H.heidelbergensis or a H. floribunda linkage (but see set B). The distribution southwards to Bredasdorp (Rooivlei and Adoonskop) and then westwards to Napier may be in an integrated state with H. floribunda (i.e. neither one species nor the other). At Napier the appearance of the plants still does tend to H. maraisii, so that we have a population that appears to be this discrete species juxtaposed with H. mirabilis var. badia. They have the same flowering time and based on the westward changes in appearance of badia, it may actually be a product of “hybridization” or “introgression” of H. mirabilis and H. maraisii. Thus a ring-cline may exist, and what appears to be two distinct species could be due to the different routes taken in space and time to a point where they grow in close proximity and are interbreeding or have interbred. The southwestern forms of H. maraisii may be a composite of this species with H. mirabilis but also involving H. heidelbergensis and H. floribunda. H. rossouwii could also come into the reckoning, except that this is a spring-flowering species like H. mutica,as opposed to late-summer flowering as is the mirabilis/maraisii complex.
(Note:- the item numbers are used to indicate map position – if space does not permit, a site can be assumed to be in close proximity to the number given in squared brackets).
- Stormsvlei MBB7257. This is from the original site shown to me by Payne where I thought the plants had been destroyed. I found about 70 plants on a much degraded site where a wattle tree had just been removed from the top of a small outcrop of Bokkeveld shale. The site is only about 50m away from the point shown to me by Payne but the shale ridge is so degraded that it is unlikely that more plants exist there. The plants were very exposed and burnt by the sun and it was only under the dense shade of a plant of Galenia africana that I found three dark-green plants that I could comfortably equate with my original opinion that it closely resembled plants from north of McGregor. From the point of view of description and typification, it is important to note from the pictures that the leaf surfaces are with spines and thus do not match von Poellnitz’ description. Not all the plants were with retused leaves either. The species Galenia africana is a vegetation pioneer and an indicator of disturbance.
- Stormsvlei MBB6954, JDV96/19. The plants illustrated are from north of the river in quarternary river boulders [1]. In situ they have strongly retused leaves and I use the word “retuse” under the impression that an original meaning of “retuse” was to indicate “bent back like a thumb”. The impression in the literature is that such retused leaves have a distinct end-area. This is not so. The leaves of many species in the southern Cape, including H. herbacea, H. reticulata, and H. rossouwii which are characterized by having rounded rosettes with incurving leaf-tips, are swollen convexly on the inner lower surface. This convexity can be emphasized to the extent that it manifests as an end-area. In addition to this variation, the leaf end may be either incurved or recurved. The original plants of H. maraisii from exposed vertical shale south of the river were strongly retuse and this character was to my knowledge maintained in the plants that were once in my possession at the Karoo garden. Plants from this site north of the river are retused in habitat but not always so in cultivation. Thus they become very similar in appearance to plants in the sandstone ridges within the pass viz. H. mirabilis var depauperata [2]
- SW Heidelberg MBB6663, JDV97/35. The plants are very similar to collections from Swellendam (10, 11) and from Rondeheuwel (14) and Verdwaalskloof (15). The habitat geology is Enon conglomerate, as is 10. 11, 14 and 15 are on Bokkeveld shale.
- Morning Star MBB7218, MBB7220, Witheuwel MBB7226, Somona MBB7229. It is difficult to clearly associate these two populations with the sets under discussion here. The problem is that while it seems obvious that H. magnifica and H. maraisii are conspecific, the truth is that east of Heidelberg H. magnifica is demonstrably continuous with H. heidelbergensis. I have expressed the opinion that 3 and 4 may also be influenced by H. floribunda.
- Andrieskraal MBB7237, MBB7238, Skeiding MBB7239. These three populations are obviously related to H. magnifica and I should mention that H. magnifica var atrofusca is a problem because its real position taxonomically may be dominant to the typical variety. Thus these Skeiding plants are closer to the var. atrofusca (with obtuse leaf-tips) than they are to the typical var magnifica (with an end bristle). The geological substrate is ferricrete as at Spitzkop, Heidelberg (6). While I have stated the relationship to H. magnifica, it must be remembered that this species should be seen as an extension of the problem being discussed in this paper. (Spitzkop is also the name for the type locality of H.magnifica var. atrofusca at Riversdale).
- Spitzkop (Heidelberg) JDV89/2. This population could also be considered to be maraisii-like but the leaf surfaces are inclined to be finely tubercled without the spine-like surface armature commoner in H. maraisii. But this is also not diagnostic, because there are magnifica-like populations where the individuals are with either finely tubercled surfaces or with very pronounced tubercles and/or surface spination. Descriptive detail is not adequate to make any distinction between the populations 5 and 6 and H. magnifica further east.
- Koppies MBB6879. Here the plants show floribunda-like character and this is more evident in the next (8). H. rossouwii is present nearby.
- S Oudekraal MBB6881. H. rossouwii is also present about 1km to the north. These plants are even more floribunda-like and I have included some of these populations in a resume of H. floribunda.
- Stuurmanskraal J.Esterhuizen in JDV93/64. This population is geographically nearer to the Breede River and to Malgas and the plants have a similarity to that south and west of Malgas.
- SW Swellendam MBB6644, MBB6860. The plants tend to have erect or sub-erect leaves. The leaf ends are generally not retused and the tips are often slender and floribunda-like.
- W Swellendam MBB6861. These Swellendam populations mirror that at Heidelberg (3) and the plant form repeats at Rondeheuwel (14) and at Verdwaalskloof (15) to the west. I have always regarded the Rondeheuwel population as indeterminate between H. mirabilis and H. maraisii and considered that it might be hybrid. However, I have with time become increasingly skeptical that hybridization accounts for problem variability. My impression is that the sequence from Heidelberg through Swellendam to Rondeheuwel and Verdwaalskloof support the contention that there is genetic continuity that is not a product of hybridization. The similarity suggests to me that this may be archetypal for the populations in the five sets that I am dealing with here.
- 12. Witkop/Uitvlugt MBB7242, MBB7243. These two populations are on ferricrete inselbergs that are a conspicuous feature of the landscape between Stormsvlei and the Potberg in the southeast. Together with the next two populations the plants link Stormsvlei to the Potberg mélange that I think may relate to H. floribunda as well as to H. heidelbergensis as I already have alluded.
- Die Kop MBB7244, MBB7245. (Not Die Kop Wydgelee MBB7500) These are south of the two former collections and are in the same mould. I treat them separately because the one collection is on a white-clay pressure burst and the plants are less scabrid than in the other collection that is on reddish pebbly soil. Both sites are on ferricrete.
- Rondeheuwel MBB6862, JDV90/32, JDV96/43. My first impression of this population about 1972 was that it was hybrid between H. mirabilis and H. maraisii. Yet even then I was expecting some king of continuity between these two “species” other than hybridization.
- Verdwaalskloof MBB7074. This site is near to Riviersonderend and is one of the strongest links between the series in Set C that could perhaps comprise a separate H. mirabilis.
- Napky MBB7030, JDV96/18. This is near the origin of the von Poellnitz H. maraisii var simplicior. It is now south and west of the Breede River and there is an element of floribunda in the plants viz. a more elongated leaf with flattening and rounding of the leaf-tip and the slight twist evident in that species (see 7 & 8, and also true for 3, 9 and 10). It should be noted that in my treatment of H. floribunda (Umdaus Press, in ms) I suggest that this may be a juvenile character perpetuated in that species.
- Tarentaal MBB 6539. One of the few populations that transgresses the tendency to a geographic drift in similarity of populations. (Some of…)The plants in this particular population are very similar to robust forms of H. magnifica var atrofusca as at Riversdale.
- Witklipkop MBB6890. This and the following four populations are problematic and further populations occurring along the lower Breede River exacerbate the situation. They need to be included here but at the same time they are clearly connected to a different set of populations that I relate to H. heidelbergensis. The leaves are smaller, more numerous, more elongate and generally without an end-area.
- Brakfontein MBB6886.
- Juliesfontein MBB6882. In my Revision I cite an early collection of mine MBB1211 from here under H. maraisii. I also included a collection of P.V. Bruyns from Ziekenhuis (see 24) because the plants were very small and seemed similar. There are good herbarium specimens to show this. However, when I re-collected at Juliesfontein after the publication of my Revision in 1999, there were no plants at my remembered site and much larger plants nearby which I would have had difficulty putting with H. maraisii. The vast mass of material I have seen since has not made a taxonomic decision any easier. I can honestly say that recognizing a taxonomic difference between these two populations (20 and 24) would serve a very limited purpose.
- N Potberg MBB6889 [20]. Chris Burgers (CB2506) first sampled this population and I also cited it under H. maraisii. This is not a comfortable solution.
- NW Potberg MBB6544, JDV96/54 [20]. H. variegata occurs here too and may infuse populations in the area. I have one record of plants that seem to suggest this, but this would need to be covered in a discussion of H. variegata itself.
- Jakobsrivier JDV92/85, JDV87/50. Needs to be considered with the following. They are very small plants with blunt and rounded leaf-tips suggestive of the Potberg collections 18 –22.
- Ziekenhuis P.V.Bruyns in JDV87/5, Uitrus MBB7250. The first is a collection that P.V Bruyns made east of the river. These are very small plants with erect leaves much as I remembered the original Juliesfontein collection of mine. They flower in January as this whole complex generally does. The geology of the east bank of the river is Bokkeveld shale. At Uitrus it is rather uncertain as while there are exposed shales in the vicinity they are overlaid by pebbly deposits of recent origin. This may be very important in relation to the juxtaposition of this site (and 22) with Set B.
- Rooivlei MBB6638. The following five populations are a continuance of the Napky (16) one where there seems to be a floribunda impact and the plants are also very dark green often conspicuously tubercled. The leaves are thick and blunt. An element also occurs here which is apparently H. heidelbergensis, and H. turgida is also present. The presence of a separate heidelbergensis-like population is confounding. Those plants flower a little earlier and should be discussed with plants from Kathoek, Beyersdal, Haarwegskloof. They cannot be excluded from a total concept of H. mirabilis that I am suggesting here, and as I am going to do for the same reason that this conspectus excludes populations from beyond Heidelberg.
- Adoonskop MBB6640. These are like the Adoonskop and Napier variants (27) and easily put with a concept of H. maraisii if it were feasible to separate such an element from H. mirabilis in the overall relationship of the populations. The plants tend to have rounded leaf-tips as is the case in the following four populations.
- NE Bredasdorp JDVsn – no specimens collected. Plants as above. This would have been an important record because of the proximity to Bredasdorp and to H. mirabilis as the var. sublineata. The site was also home to Aloe brevifolia and sadly the site is so degraded that neither Haworthia nor Aloe can now be found there. MBB7334 is a very recent collection, nearer o Bredasdorp and plants are very similar to (29)
- Napier MBB7259, JDV85/15. There is a substrate difference in that at this site the soil is pebbly quartzitic whereas at the following site nearby (29) it is finely slabbed, coarse grained sandstone. MBB7259 is about 4km north and the substrate there is true micaceous shale.
- Napier JDV85/96, MBB7247 [28]. Whereas plants at the former sites are blackish-green, here the plants are reddish-tinged.

A01 MBB7257.1 Stormsvlei 


A02 JDV96-19.1 N Stormsvlei 


A03 JDV97-35.1 SW Heidelberg 
A03 MBB6663.1 SW Heidelberg 


A04 MBB7218.1 Morning Star 


A04 MBB7220.1 Duiwenhoks 

A04 MBB7226.1 Witheuwel 

A04 MBB7229.1 Somona

A05 MBB7237.1 Andrieskraal 

A05 MBB7239 Skeiding 
A06 JDV89-2.2 Spitzkop, Heidelberg 


A07 MBB6879.1 Koppies 



A08 MBB6881.1 S Oudekraal 





A09 JDV93-64.1 Stuurmanskraal

A10 MBB6644.1 SW Swellendam 




A12 MBB7242.1 Witkop 

A12 MBB7243.1 Uitvlugt 






A13 MBB7244.1 SE Die Kop 



A14 JDV90-32.1 Rondeheuwel 
A14 MBB6862.1 Rondeheuwel 


A15 MBB7074.2 Verdwaalkloof 





A16 JDV96-18.1 Napky 
A16 MBB7030.1 Napky

A17 MBB6539.1 Tarentaal 




A18 MBB6890.1 Witklipkop 

A19 MBB6886.1 Brakfontein 




A20 MBB6882.1 Juliesfontein 


A21 MBB6889.1 N Potberg 
A22 JDV96-54.1 NW Potberg 

A22 MBB6544.1 NW Potberg 
A23 JDV92-85.1 Jakobsriver 


A24 JDV87-5.1 Ziekenhuis 


A24 MBB7250.1 Uitrus 




A25 MBB6638.1 Rooivlei 

A27 MBB7334.2 H. maraisii. NE Bredasdorp. 



A28 JDV85-15.1 N Napier 
A28 MBB7259.1 Napier Station 






A29 JDV85-96.1 NE Napier 
A29 MBB7247.1 H. maraisii. NE Napier. 






B. Haworthia mirabilis towards the Potberg:-
This set is actually anomalous and consists of only three populations associated with Table Mountain Sandstone and calcareous limestone. At Karsriver, a short distance from 27 is where one finds H. rossouwii var petrophila (which I had previously allied to H. variegata) in limestone. Further east is H. variegata var modesta and to the south of that H. rossouwii var calcarea (previously as a variety of H. mirabilis). It is obvious that the classification is difficult and dubious. However, C Burgers made another surprising collection (CB2018) from Buffelsfontein [CB] south of the Potberg. I cited it in my revision under H. mirabilis var calcarea. I have had a problem with a specimen collected by Prof. H. Compton cited “from the Potberg sandy flats”. That specimen is of a retusa-like plant that I thought might perhaps be representing either H. mutica or H. mirabilis but I did not cite the specimen in my revision because I was so doubtful about it. I thought this could have been what Burgers found, but his collection was in rock crevices and not on the sandy flats. However, Adam Harrower made a collection (ADH594) from 20km further east, also in rock crevices and clearly the same as the Burger collection. On a very recent trip my wife and I found a similar plant (31 MBB7248) on the northeast end of the Potberg that I think may fit here. The problem is its proximity to 24 only 3km away.
(See Brutality of reality in Haworthia).
- Sandhoogte ADH594, MBB7251. The site is a most unprepossessing and unlikely looking one from a distance. The vegetation is dense waist-high fynbos, and Adam Harrower found the plants in the rocky north-facing border to a stream. They can be compared with H. turgida var longibracteata (except they are less proliferous) and also with H. mirabilis var sublineata. There is also a likeness to forms of H. rossouwii var calcarea, albeit distant. More striking is a curious resemblance to H. magnifica var splendens from near the Gouritz River Bridge.
- Ballyfar MBB7248, MBB7249 [off map E of 22}. These two populations are only a few kilometers from 23 and 24, but they are on sandstone. What is more is that the plants are larger but have fewer leaves that are strongly retused as in the Compton specimen. What is striking about the plants is the presence of numerous surface spines and one is forced to consider the resemblance to a population of H. magnifica east of Riversdale with similar spination and general appearance, only proliferous. The site is about 3km due east of the Sandhoogte population and this suggests that the two populations are of the same species. There is an interesting dimension to the problem mirrored in the preceding (30). This is the occurrence of H. turgida at Diepkloof only 15km to the northwest and only five kilometers from Burgers 2018 at Buffelsfontein across the Potberg Mountain. A decisive factor may be flowering time, because H. turgida flowers in spring and the other populations considered here flower in mid-summer. But the main point is that the relationships are complex and varied.

B30 MBB7251.1 Sandhoogte De Hoop 




B31 MBB7248.3 Ballyfar 


C. Haworthia mirabilis and its variation south of the Riviersonderend Mountains:-
This set also starts with Stormsvlei and is the south and westward set which is almost exclusively mirabilis. There is a possibility of a connection across the Riviersonderend Mountains closer to Villiersdorp where it may be suggested that H. maculata is implicated too, but this will instead be considered in Set 3. These are plants with a tendency to redden. They are usually larger than those in set 1. The height may be as much as 100cm and rosette diameter 110mm.
- Stormsvlei Pass MBB6955. This population is in sandstone and immediately there is a significant difference to the plants immediately south (1) and north (2) of the Riviersonderend River a short distance to the south. There is no evidence of an end-area to the leaves. The leaves are longer and more slender and they show the redder colour one associates with H. mirabilis elsewhere. Some of the plants are very highly spotted on the back or under surfaces. This is a character that has been associated with H. mirabilis when in fact quite common to find plants with or without such marking in many of the populations in these sets.
- E Stormsvlei Pass E. Aslander sn. MBB7252, MBB7253 and MBB7254. In exploring the southern slopes of the Bromberg I came across several populations. The geology here is rather complex as it is the close interface of sandstone and shale, with the pebbly patches associated with ferricrete (Gibbaeum austricolum is present). At one site (MBB7254) the plants were small and with narrow elongate leaves on a coarse-grained sandstone ridge, while immediately north on a pebbly band there were plants with fewer, shorter and broader leaves. There is no difficulty in allying these plants with 32.
- Greyton JDV96/47. The continuity westwards is disrupted by lack of records and there does not seem to be suitable habitat between the Riviersonderend River and the mountains. A population at Lindashof recorded by G.G. Smith appears to have been destroyed in a road-making operation. This population at Greyton is of rather small plants with conspicuously finer and intense spination.
- Uitkyk MBB7092, JDV95/97. This is in the extreme west for the set, at Elandskloof west of Genadendal. It is a very rocky south-facing site and the plants are robust and proliferous.
- Leeukop JDV90/34, JDV85/98. This is the locality for the var rubrodentata where the plants have long slender leaves with intensely red spines – hence the name.
- Genadendal JDV90/33.
- Skuitsberg JDV88/70, JDV96/101. Here there is quite an extensive population with a similar one also present on the adjoining farm Nethercourt. The plants are generally very robust and were described originally as H. emelyae var beukmannii.
- Jongensklip MBB7059. The interesting thing about this population is that it fills somewhat of a void between populations known respectively north and south of the Caledon/Bredasdorp wheatlands.
- Goudini MBB6537. This population is the southwestern-most for the species. I have grown it from seed and the variability among the seedlings is quite striking, which confirms my opinion that the taxa described from north of Napier are not significant as discrete varieties.
- Schietpad MBB4642. JDV94/90. See 43.
- Fairfield MBB6643. See 43, except that the plants are very proliferous.
- Mierkraal (Napier) MBB7091, S Mierkraal MBB7260. The plants now constitute a general form that appears in the preceding three populations and there are several such populations in the area between Mierkraal and Fairfield (42) and I do know that I have seen it in the Klipdale area to the northeast. The varieties H. triebneriana vars multituberculata, napierensis (Schietpad, 41) and turgida, originated north of Napier and yet I would not consider the variability to be population based nor even so significant in terms of the variability of H. mirabilis var triebneriana as I would designate the taxon. MBB7259 is a population south of Mierkraal, the plants tend to have more elongate and erect leaves. The site is geographically nearer to 40 (MBB6987) which is H. mirabilis var badia.
- NW Napier MBB6635. Although the geology is the Bokkeveld group, the rock here is very quartzitic and the vegetation alters from Cape Fynos to renosterveld over a very short distance. The site is that of H. mirabilis var badia. The geology in this area is quite complex and there is also a small ferricrete inselberg to the east.
- W Napier MBB6987. This population is a western continuation of the previous one and is also the var badia. However, there is a tendency for the leaves to be more erect and narrower. I do speculate that this var badia may have arisen from an interaction of H. mirabilis and H. maraisii. The four populations (28 & 29 of set A, 44 & 45) flower and seed at the same time, seedlings of both are highly variable and both have companion populations which suggest the deduction of continuity across a very strong geological but weak breeding barrier.
- Bredasdorp N JDV86/8, MBBsn, MBB7261sn. This locality is immediately north of the town on the riverbank. The area was highly invaded by alien Acacia and the haworthia is no longer there. The plants are somewhat similar to those south of the town, but more robust and with more strongly retused leaves. This is a close approach to H. maraisii (26) and may parallel the situation at Napier (28 & 29) where this species and H. mirabilis var badia (44 & 45) are almost sympatric. My MBB7261 are a few plants rescues from about 300m northwest of JDV86/8 from a site overgrown with bluegum. These plants had erect leaves.
- Bredasdorp S MBB6639. The site is a north-facing sandstone mountain slope. The plants are highly variable and are named as H. mirabilis var sublineata [47]. The leaves are long and slender. Some clones are proliferous and others not.
- Mierkraal (Bredasdorp) MBB7090. H. mundula was described from this locality which is unusual in being recent sandstone and on the very southern fringe of the distribution of H. mirabilis. The plants are very proliferous and their leaves have a very pronounced end-area that is also smooth. The population is thus fairly different from others and this is unfortunate because it is in my opinion the source of the type and hence is the var mirabilis. I first saw the population in 1969 and can testify to significant changes in the precise whereabouts of large proliferous mats in the past, as opposed to more scattered less proliferous clumps in the present.

C32 MBB6955.1 Stormsvlei Pass 







C33 MBB7252.1 Noukloof 
C33 MBB7254.1 E Stormsvlei 
C34 JDV96-47.1 S Greyton 
C35 JDV97-94.1 Uitkyk 
C35 MBB7092.1 Uitkyk 



C36 JDV85-98.1 S Greyton 



C37 JDV90-33.1 W Genadendal 



C38 JDV88-70 Skuitsberg 
C38 JDV96-101 Skuitsberg 
C39 MBB7059.1 Jongensklip 


C40 MBB6537.1 Goudinie 






C41 JDV94-90 Schietpad 
C41 MBB4642 Schietpad

C42 MBB6643.1 Fairfield 




C43 MBB7091.1 Mierkraal, Napier 







C44 MBB6635.5 NW Napier

C45 MBB6987.1 W Napier 

C46 JDV86-8.1 N Bredasdorp 

C46 MBB7261.1 NW Bredasdorp 


C47 MBB6639.1 S Bredasdorp 




C48 MBB7090.1 Mierkraal, Bredasdorp 

D. Populations north of the Riviersonderend Mountains:-
Stormsvlei is where the Riviersonderend Mountains begin to tail away to the lowland wheatfields. The Riversonderend River cuts through the mountain about 10km east of Stormsvlei, and the Stormsvlei Pass itself is a rift through the mountain tail that allows road access from north to south. The mountain to the west is Table Mountain Sandstone. Rainfall is high and the vegetation is Cape fynbos. It must be a formidable barrier to genetic flow and unsuitable habitat for succulent plants. There are very strong geological gradients coupled with precipitation differences north and south of the mountain. The geological gradients also have an east-west pattern as opposed to the essentially north-south pattern south of the mountain. Further to the west, folding and faulting have resulted in a huge wedge of more recent formations appearing between Worcester and Villiersdorp. McGregor is virtually at the southern end of a wedge of older Bokkeveld formations that run northwestward to Robertson. There it meets the great Worcester fault line and a massive igneous pluton. Rocks of the older Malmesbury Supergroup and rocks of the much younger Karoo Supergroup are juxtaposed with igneous intrusive rock. The plants dance to this pattern and this is reflected in the differences found in Haworthia populations. This set comprises the populations on the Riviersonderend Mountains itself and along the northern foothills. The plants are intermediate between sets C and D and include significant ecotypes, as for example the Boesmanspad plants that resemble plants in the H. turgida complex, the Moddergat plants associated with H. maculata and the Trappieskraalkloof plants which are the maraisii-like element.
- NE Stormsvlei Pass (Brakfontein) MBB7087. This is on a dry north-facing slope where the vegetation is Fynbos and close to the transition to Renosterbos. The geology seems to be recent scree and sand. The plants have erect to sub-erect leaves, are quite narrow and have strong marginal spination. They thus seem to equate with plants within the Pass (32) in quartzitic sandstone of the Table Mountain Group and seem to differ from plants close-by to the northwest (67, MBB7086) that are on a shale/sandstone formation of the Bokkeveld Group.
- Boesmansrivier MBB6711. The Boesmansrivier Valley is a very long and deep incision into the mountain running southwest to northeast. The plants are similar to Stormsvlei Pass (32) but the undersurfaces are often more strongly spotted and the faces of the leaves are frequently also very attractively spotted with translucent dots.
- E Olifantsdoornkloof MBB6686. The habitat here is slightly drier and more exposed than the preceding and the plants have shorter squatter leaves.
- Olifantsdoornkloof MBB6685 [51]. At this site there is again a deep incision into the mountain and the habitat is more sheltered and moister. The plants have longer leaves. Von Poellnitz named H. triebeneriana var diversicolor from this population and the name refers to his observation “plants therefore appear diseased and no plant agrees fully in colour with the other”. It is instructive to think what he would have thought if he had seen plants from the preceding populations extending through to the following 15 in this set.
- Hoeksrivier MBB6575. What is surprising and different about this population, also in a rift into the mountain, is that the plants have the rosette shape of H. herbacea. Although the leaves have the swollen mid-section, the ends are curved inward so that the rosette is more rounded. These plants are particularly “diseased” as in 52.
- Houtbaaiskloof MBB6760, JDV97/163, MBB7241. Houtbaaiskloof is a very deep incision into the mountain immediately south of McGregor, which meets a geological fault line north of the Hoekrivier. So I was able to make collections both north and south in this pseudo-gorge. This is where von Poellnitz’ H. whitesloaneana originated The leaves revert to the tendency to have an end-area but the “diseased” appearance is reminiscent of 52.
- Takkap MBB6723, JDV89/63. In this incision of the Takkap River into the mountain the plants become increasingly mottled and also slightly softer in texture coupled with increasing leaf numbers although they are still small in size.
- Dwarswaterkloof MBB6865, JDV89/67. As with the Takkap plants (55) the plants are greener and quite proliferous. The Dwarwaterkloof is again a very long and deep incision into the higher peaks of the Riviersonderend Mountains. It is not surprising that the plants have continuity with the following populations.
- Boesmanspad JDV84/67, JDV88/63, JDV89/67 . The Boesmanspad refers to a footpath that traverses the Riviersonderend from McGregor to Greyton. There is a deep valley called Boesmanskloof that runs from Greyton in the south towards McGregor. It is a high rainfall area and it is quite surprising to find Haworthia in this gorge. Furthermore the plants are small and very turgid so that the comparison can be fairly drawn with that of plants of H. turgida var turgida from similar wet and rocky situations in the Langeberg mountains e.g. Southey’s Pass or Garcia near Riversdale. The flowering time and flower itself in the context of distribution and flow in similarity suggests that this must be an ecotype related to populations both north and south of the mountain range.
- Koeniesriver MBB6714 [56]. These are in a more exposed situation than the previous two and with the following (59) are again more reminiscent of plants from the Stormsvlei Pass itself (32).
- Rietvleikloof MBB6720. The topography of this mountain incision is interesting because it is practically the last of a north/south series. The next incision is a major faultline that runs southwest/northeast. This forms a valley largely in the lee of high mountain (Jonaskop).
- Moddergat JDV96/34, MBB7270. This population needs to be carefully considered. It oversteps the distribution of H. herbacea and the var. livida that I described from just east at Lemoenpoort. I also described H. herbacea var lupula from an improbable habitat deep in the mountains east of Villiersdorp. I associate the Moddergat plants with H. maculata to the north at Brandvlei and this has still further associations in the Langeberg Mountains. However, the flowering time and appearance of the plants do suggest that a connection with H. mirabilis has to be considered even if improbable. I have indicated already that this area is geologically diverse.
- Trappieskraalkloof MBB1210, Droeriviersberge MBB7263, MBB7264, Haumanskloof MBB7265. The first I considered from memory to be a very significant population that could better be placed in Set E. The plants are small, greyish in colour. The rosette is somewhat globular with short retused end-areas and very similar to 65. I initially made this collection MBB1210 in my first few years at Worcester, and it is only at the close of this work that I have revisited the area. The area is difficult to access and is very dry. The terrain is extremely rocky and rugged. Dywka Tillite interfaces with Witteberg Quartzite and the vegetation can change dramatically over a distance of a few meters. It is not clear how the latter three populations relate to the traditional perceptions of H. maraisii and H. mirabilis as discrete species, and nor to their relation with H. maculata, which is not far to the west at Moddergat (see 60 JDV96/34. MBB7270). It is evident that they are connective between sets D and E.
- Whipstock MBB6864. Again one has to take geology into account as I discussed for the set. At Moddergat and Trappieskraalkloof one is confronted with more recent geological phenomena. Whipstock is at the southern end of a similar quartzitic formation and the plants follow the trend indicated in the early items of this set. The plants are somewhat mottled and an end-area is often apparent.
- Klipbergdam MBB6681, MBB6682, JDV97/48. These plants demonstrate continuity between the early members of this set and the following set.
- W McGregor MBB6673, JDV96/49. This is in the McGregor Nature Reserve and is the source of the original H schuldtiana as was conveyed to me by G.J. Payne.
- N McGregor MBB6646 [64]. This population is the origin of the names H, schuldtiana vars minor, subtuberculata and unilineata. Obviously the population seems to be unusually variable because of the varieties that it has given rise to. But this is not so and simply reflects the weak taxonomic precepts of the time which seem to be so difficult to dispel in the present.
- SE McGregor MBB6687 [64]. The plant I illustrate appears to have very pronounced end-areas to the leaves. But I have not explored the area thoroughly as the plant fits conformably with the populations I am listing.
- NW Stormsvlei Pass MBB7086, & Vaandrigdrift MBB6534, JDV96/25. I have often observed plants northwest of Stormsvlei Pass and simply accepted that they belonged to the discrete species I had conceptualized viz H, maraisii. They now easily fit the conceptual model that the elements around Stormsvlei and the Pass are actually one and the same species. I have not collected at the latter two sites further to he east. Plants I observed there would have fitted those illustrated in 61, 64, 65 and 66 as well as with many in set E. This would correspond to a concept of the element H. maraisii as I recognized it in my Revision.
(See A population of H. heidelbergensis and note MBB7513=6534)

D49 MBB7087.1 NE Stormsvlei 



D50 MBB6711.1 Boesmansrivier 





D51 MBB6686.1 E Olifantsdoornkloof 





D52 MBB6685.1 Olifantsdoornkloof 



D53 MBB6575.2 Hoeksrivier 


D54 JDV97-153.1 Houtbaaiskloof 


D54 MBB7241.1 N Houtbaaiskloof 




D55 MBB6723.3 Takkap 


D56 MBB6865.1 Dwarswater 





D57 JDV84-67.1 Boesmanspad 



D58 MBB6714.1 Koeniesriver 




D59 MBB6720.1 Rietvleikloof 



D60 JDV96-34.1 Moddergat 


D60 MBB7270.1 Moddergat 

D61 MBB7263.1 Droerivierberg 



D61 MBB7266.2 Droerivierberg 



D62 MBB6864.1 Whipstock 





D63 JDV97-38.1 Klipbergdam 





D64 JDV97-49.1 W McGregor 



D65 MBB6646.1 N McGregor 





D66 MBB6687.1 SE McGregor 

D67 MBB7086.1 N Stormsvlei Pass 
E. Haworthia maraisii north of Riviersonderend Mountains:-
This is the Worcester/Robertson Karoo set that I have simplified by excluding H. maraisii var notabilis. The reason I have done so is because of a manuscript of mine dealing with the epithet intermedia which is with the Editor of Haworthiad. That manuscript deals with the impact of the granitic pluton at Robertson and associated dolomite on Haworthia although I have there not really discussed the geology as such. At Klaasvoogds where there is also a dramatic shift from ancient to very recent rocks and replication of the problem of a var. notabilis that is not dealt with in this paper. This set illustrates the populations that I used to formulate the concept of a species H. maraisii. It also includes the “escape” into the Little Karoo and the possible eventual link with H. emelyae east of Barrydale. The plants are characteristically very dark coloured and perhaps smaller in comparison to sets A, B and C. However, I think I can demonstrate reversion to a mirabilis-like plant and also similarity to H. heidelbergensis. I also include two populations that suggest a more far-reaching link to H. emelyae north and east of Garcia Pass, north of Riversdale.
- Bosfontein JDV97/58. These are plants in a continuum from Trappieskraalkloof (61) on the same geological formation viz. Witteberg Group rocks.
- Dublin F.J.Stayner in KG400/61, JDV94/18. The significance of these plants is that they were particularly dark-green, very tuberculate and the leaves were erect. Although I saw plants at the site with Frank Stayner I have not been able to find them at the site since. Plants do occur again a short distance to the east, but they do not have the same very erect and narrow leaves of Stayner’s collection. A plant is illustrated in my revision where it is under the number JDV94/18 and attributed to Roy Littlewood who was instrumental in first drawing attention to it.
- Rooiberg MBB6956. This is north of the Breede River from the previous locality listed. The Rooiberg is quite a substantial massif and houses what can be regarded as discrete populations. Included in these is the very curious H. herbacea var flaccida. I only observed five plants of that and suspect that it is of hybrid origin with the putative parents being H. maraisii and H. reticulata rather than H. herbacea.
- Agtervink MBB6647. The following eight populations illustrate the geographic core of my concept of an H. maraisii.
- N Agtervink MBB7176 [72].
- Sandberg MBB7174, JDV98/18.
- Nekkies JDV85/89, JDV96/24, EA083, EA1408. This population is very close to the Robertson pluton and I did associate this site with transition to the var notabilis. I doubt if that this is actually true for this local area. Etwin Aslander made a collection (EA1408) from very close to Wolfkloof where the notabilis complex occurs, and this maraisii variant could be compared to those in 80 north of Ashton and even with 32 in Stormsvlei Pass.
- S Sandberg MBB6670, JDV97/38.
- Koningsriver MBB6672.
- Vrolikheid MBB6968, W Kanonkop MBB6967, E Witkranz MBB7077. These and the next three populations are to the west of an area that should contain plants of the interface I would like to illustrate. However, it is a very dry area and not visitor friendly.
- Steenbokshoogte MBB6966, Klein Spitzkop MBB6969.
- Grootrivier MBB6677.
- Breede River Bridge (Robertson) MBB6648. I noted this population in my Revision with the comment that “note can be made of forms near Robertson which are rather similar to H. pubescens. The plants tend to have the same globular rosette shape and also the pubescence. However, the flower and flowering time is typical of H. maraisii as I defined it.
- Muiskraalkop MBB6961, JDV87/208. This is the type locality for von Poellnitz’ H. schuldtiana vars robertsonensis as indicated to me by Payne. However, von Poellnitz does cite more than one locality for each in his description. The plants do have more slender erect leaves than most in this set but the distinction gets lost the more material that is considered.
- Rietvlei MBB7055. This population is an odd one that I used in developing a justification for treating the element notabilis as a variant of H. maraisii. I was looking at the plants on a south slope under bush whereas it transpires that the bulk of the population is higher on the ridge in Enon conglomerate. Plants further north on Peninsula sandstone (Kranskop) are quite different and this difference compounds to the northwest where the plants are on Malmesbury shale.
- Ashton MBB6668. This site is also Enon conglomerate and the plants are quite large and the leaves are rather recurved than with an end-area. The surfaces also tend to be smooth. Hence they do appear somewhat mirabilis-like cf. 32.
- Cogmanskloof MBB6875 . These plants can also have leaves quite strongly recurved and there is a close similarity with the following.
- Baden, Montagu JDV97/55, EA068.
- E Montagu PVB7588. Malherbe of Sheilam stated that he had seen “small black plants” in the Koo valley north of the Langeberg. I have not been able to confirm this other than seeing them immediately northwest of Montagu. P.V. Bruyns recorded the species about 20km to the northeast (Dobbelaarskloof) and also at this site due east of Montagu. The plants have highly retused leaves but do not deviate substantially from others in this set.
- Barrydale MBB6667, JDV97/39, MBB7214. J Dekenah knew these plants at Barrydale and it is surprising that some varietal name or other did not evidence this. It is difficult to interpret outside of these sets and a complication is a very odd population south of Southey’s pass (MBB6666) that I have perforce regarded as H. magnifica var magnifica. It is an embarrassment to me because it has resemblances to H. retusa and to H. mirabilis as much as to H. magnifica. In fact plants from there do not closely resemble H. magnifica in the form that it is manifest in 5 and 6 of set A. It (87) is important because of the following item.
- Kleindoringrivier GDMarx in MBB6741. This population is midway between Barrydale and Muiskraal where H. emelyae var major occurs, and it appears to be a connecting link. The flower and flowering time links it to the west and to these sets.
- Loerkop MBB6979. This is southeast of Ashton and the plants have retused end-areas.
- Viljoensdrift MBB6518. This is at the foot of the massif Elandsberg and seems to be in the continuum with 74, 75 and 76.
- Middelplaas MBB7079. This population is similarly in some kind of juxtaposition between this set and the next and I interpret it like that in view of unlisted populations I have seen in this area.

E71 MBB6647.1 Agtervink 






E72 MBB7176.1 N Agtervink 





E73 JDV98-18 Sandberg 
E73 MBB7174 Sandberg

E74 EA083.1 Tierberg 

E74 EA1408.1 NW Robertson 

E74 JDV85-89.1 Die Nekkies 

E74 JDV96-24.1 Die Nekkies 
E75 JDV97-38.1 SW Sandberg 
E76 MBB6672.1 Koningsrivier 


E77 MBB6967.1 W Kanonkop 


E77 MBB6968.1 Vrolikheid 





E77 MBB7077.1 E Witkranz 


E78 MBB6966.1 Steenbokshoogte 



E79 MBB6677.1 Grootrivier 

E80 MBB6648.1 Breede River Bridge 

E81 JDV87-208.1 Muiskraalkop 
E81 MBB6961.1 Muiskraalkop 



E82 MBB7055.1 Rietvlei 


E83 MBB6883.1 N Ashton 





E84 MBB6875.1 Cogmanskloof 

E85 EA068.1 Montagu 



E86 PVB7588.1 E Montagu 




E87 JDV97-32.1 Barrydale 

E88 MBB6741.1 Kleindoringrivier 


E89 MBB6979.1 Loerkop 

E90 MBB6518.1 Viljoensdrift 



E91 MBB7079.1 Middelplaas 


F. The interplay of Haworthia maraisii and H. heidelbergensis:-
This set includes the var. meiringii at Bonnievale and a set of populations that suggest a closure with Set A and the emergence again of H. heidelbergensis. Many of the collections listed here seem to be entirely new to the literature of the genus. The tendency in this group, as a whole, is to erect leaves. They are also small with a maximum height of 50-70cm and a rosette diameter of 20-40cm. The geology of the area does not seem to be excessively complex or vary substantially from the previous set except that the topography is more uniform. However, the plants seem unable to hold the profile of the previous set and there is a drift to H. heidelbergensis.
- W Goudmyn MBB6516. I start this series in the mid-west of the area because it has the more conventional forms of H. maraisii to both the north and the south. This population is west of Olifantskrans and extends on a long sharp ridge to the summit of the Elandsberg to the west. The plants are very small and have many small slender erect leaves.
- Olifantskranz, E Goudmyn MBB6676, MBB7081. I have four collections from this general area and the plants are continuous with the previous set. The leaves are a little more substantial and often very spotted. The continuity extends both east- and northeastwards.
- Olive Grove/Wakkerstroom MBB7081, MBB7084. This is the eastward extension and the plants are approaching H. maraisii var meiringii at 95. The leaves are slender and erect and relatively opaque.
- W Bonnievale MBB6509. There is also a record from north of this point which are less spinose and translucent than this particular population although the distance separating them in less than 100m. Here specifically, I find plants which I would compare favourably with H, heidelbergensis var toonensis at Matjestoon east of Swellendam. The continuity is in fact with H. maraisii var meiringii below.
- E Bonnievale MBB7085, Edendale MBB7269. These plants are very proliferous and spinose. The rosettes are globular. The var meiringii was actually based on the population to the near east of this one (MBB7269) where the vegetative similarity to H. herbacea was even more significant.
- Bobbejaanskloof MBB7082, N Goudmyn MBB7083. Northeast of Olifantskranz (93) the continuity extends, not as expected to the conventional H. maraisii as at Loerkop, but to plants here with more slender erect leaves suggesting H. heidelbergensis.
- Zandvliet MBB6964, MBB6965. Here there are two distinct populations that I include in the var meiringii together with the previous 6 populations. Plants do vary from individuals with stubby nearly terete leaves to plants with the conventional leaved wider than thick and narrowing to the apex.
- SE Loerkop MBB6981. This is a very substantial population and while it would fit in the mélange of the previous item, it starts to relate uncomfortably to the next.
- Tonteldoos MBB6974, MBB6977. These and the following to 111 are a nondescript assemblage of plants that do not relate comfortably with any of the other sets and neither do they collectively suggest any kind of taxonomic unity.
- Mardouw MBB6990. Bob Kent also has two collections from between this and the following sites.
- Nooitgedacht MBB6988, MBB6989 [102]. The first population is also a very substantial one close to eucalypts as is the following. However, the second is in natural vegetation.
- Langverwacht MBB6992. This is a remarkable population of small very proliferous plants with quite turgid stumpy leaves. The site is notable as a quartzitic inselberg marked by the occurrence of several large gum trees. The haworthia seem to be the only plants tolerating this alien vegetation as the ground is otherwise devoid of plant growth.
- Jakkalskop MBB6971.
- Witkop (Drew) MBB7036.
- NW Sanddrift MBB7056, W Sanddrift MBB7057.
- Sanddrift MBB6625, JDV89/8, JDV90/31. Although very similar to the next, this population tends to have leaves with a retuse and opaque end-area. It is also small and very proliferous but nevertheless suggests an affinity with H, maraisii as in Set D.
- N Sanddrift MBB6626, JDV87/193. This particular population comprises small plants with spreading leaves that are quite conspicuously translucent. Thus there is no difficulty in suggesting that they belong with H, heidelbergensis as does the next.
- Middelrivier MBB7037.
- Leeurivier JDV87/4. This particular population has been linked to H. heidelbergensis var scabra and the following recent collection seems to confirm that this is the continuity that should be sought.
- Nuutbegin MBB7099. This population is now south of the Breede River again and the problem is that it does not seem to relate to the initial populations in set A, but rather to those south in the Potberg area that I have not included in this particular discussion. Those populations are dealt with in a review I wrote of the H. floribunda complex and could initiate an overview of H. heidelbergensis.

F92 MBB6516.1 W Goudmyn 


F93 MBB6676.1 Olifantskrans 



F93 MBB7081.1 E Goudmyn 


F94 MBB7080.1 Wakkerstroom 


F95 MBB6509.1 W Bonnievale 


096 MBB7085.1 E Bonnievale 


F97 MBB7082.2 Bobbejaanskloof 
F97 MBB7083.1 N Goudmyn 



F98 MBB6964.1 E Zandvliet 



F98 MBB6965.1 Zandvliet 



F99 MBB6981.1 SE Loerkop 




F100 MBB6974.1Tonteldoos 




F100 MBB6977.1 Tonteldoos 



F101 MBB6990.1 Mardouw 


F102 MBB6988.1 Nooitgedacht 




F102 MBB6989.1 SE Nooitgedacht 




F103 MBB6992.1 Langverwacht 








F104 MBB6971.1 Jakkalskop 



F105 MBB7036.1 Witkop, Drew 


F106 MBB7057.1 W Sanddrift 




F107 JDV89/8 Sanddrift 
F107 JDV90/31 Sanddrift 
F107 JDV96/25 SE Drew 

F107 MBB6625.1 Sanddrift 

F108 JDV87/193 N Sanddrift 

F108 MBB6626.01 N Sanddrift 











F109 MBB7037.1 Middelrivier 




F110 JDV87/4 Leeurivier 



Discussion:-
It is wishful thinking to even hope that some taxonomic decision can flow from the above discussion that will match the traditional classification of haworthias. I consider that I set out to describe H. maraisii in the same format as a series of 5 articles that I wrote for the English Cactus and Succulent Journal entitled rather grandiosely as “Natural variation and species delimitation in the genus Haworthia” (ca 1972). I never completed the article because I could see that the 20 or so populations that I had then sampled held insurmountable problems. Since then an obstacle for me has been the seeming intellectual intransigence of readers, expressed by other writers, who like myself seem to have been wishing for some magical formula whereby the genus would unfold and disclose the obvious and comfortable units that we all wish species were – and also by lack of full exploration. My uncomfortable role has been to try vainly to educate and instruct a small willful segment in both academic and non-academic rank, that this Holy Grail is based on not only false hopes, but on false premises.
Classification of Haworthia, and indeed I contend that this will apply to many other genera, simply will not be accomplished unless there is agreement on purpose and definition. My botanist acquaintances seem to insist that there are many definitions for the word “species”. My non-botanist acquaintances are justifiably unconcerned about definition at all while they have names that they can use. That these names as latin binomials should have a particular meaning in the whole context of biology seems to be lost to both groups. Thus the nomenclatural system as expressed by the International Code serves no true biological purpose. It simply regulates and legislates for the methodology behind a nomenclatural system whether this is for true “species” or for elements recognized out of sheer lack of sampling and poor definition.
My observation derived from the material discussed in this manuscript is that I can no longer see a clear distinction between H. maraisii and H. mirabilis as the names have been transfixed by typification either by myself or by I. Breuer or by the way in which I have applied them. The problem is not simplified by the fact that at both Bredasdorp and Napier it appears that two elements are in close geographic proximity. This rather suggests that a ring-cline may exist. I suggested in my Revision, that a taxonomic solution could be possible for species (populations) together in one locality (or close proximity), but wrong elsewhere. This observation seems to apply here. Similarly I cannot see that the set of populations in E above (H. heidelbergensis?) can be circumscribed as a species in anyway that H. heidelbergensis can truly be circumscribed and seen to be discrete. More discomfort extends from the interplay of H. heidelbergensis, H. magnifica, even H. floribunda, and the complexity of that element in the east toward Riversdale and Albertinia. The relationship of this aforementioned complex to H. emelyae in the Little Karoo is compounded by the connection via Barrydale of the greater H. mirabilis set discussed in this work. There are thus other sets of species which need to be considered in the same way as has been done in this work
What I do suggest is that species are fractal and that there will never be clear focus on all life forms. There are species that are presently in sharp delimitation and species in transitional mode that are not. This is particularly evident within Haworthia where some species are quite obvious and discrete and others are not. The scale of the taxa generates the same problems as it does in the artificial rank of genera. This is that there may be one or more very distinct elements that are quite disproportionate in numbers and populations to amorphous elements. The decision in Haworthia should be reached by consensus among individuals who indeed have the wisdom, competence and experience to make sensible choices. The evidence for these qualities has so far been sadly lacking.
Conclusion:-
The set of populations that I have examined here, suggests that the probability that they can be proven to constitute two separable elements viz. Haworthia mirabilis and H. maraisii, is remote. From a practical point of view it is possible to uphold the names in the traditional way followed in my Revision, but only if it is accepted that these names cannot be applied with certainty to the greater proportion of the populations. From a botanical point of view it is patently evident that we are dealing with one species, Haworthia mirabilis. There is a greater problem in how this one species known by the populations discussed in this paper relates to the populations geographically further east. The implications of such a relationship for classifications of Haworthia may be profound and it could impact on classification of many other genera too. The obvious solution is to decisively abandon the dream of a classic nomenclatural solution to the problem. We could simply recognize that variants, which exist within the scope of any species, can be accommodated by simple descriptive statement and referred to by collecting reference and locality.
Acknowledgement:-
I have been so privileged to have made the acquaintance of many people who have facilitated my excursions. Practically every spot on the map is associated with several persons. I truly regret that it is simply impractical to name them all and I sincerely hope that my appreciation and gratitude will not be taken to be lacking by dint of omission.