
- Table of Contents
- Van Reenens Crest
- Aloe pumila, Haworthia pumila; what or who is confused?
- Haworthia limifolia – a conundrum or a lesson?
- Haworthia marxii and H. truteriorum in relation to rational classification.
- Very brief note re Haworthia nortieri flowers.
- The reality of Drosanthemum micans L.
- New finds in Haworthia.
- What do collectors need?
- Righting some misconceptions.
- Comments on H. marxii
- Haworthia maculata ↔ Haworthia pubescens, MBB8002 Cilmor.
- Addendum Haworthia pubescens MBB8011, SW Sandberg
- Genetic history – the place in Haworthia classification.
- Haworthia mutica (groenewaldii) and its twisted leaves.
- Comprehension and significance
- Habitats
- MBB6694 Kanetvlei, Hex River Valley as a variant of Haworthia nortieri.
- Haworthia minima and pumila flowers
- MBB7989 Haworthia pumila, Lemoenpoort
- Leaf arrangement in Aloe striatula
- Just what do we do with names for Haworthia?
- The absurdity of taxonomy and nomenclature?
- Variable chloracantha
- Leaf sequence in Haworthia emelyae ‘comptoniana’ over a long period
- What do you think this is?
- Kaboega
- Habitat Views
- Haworthia Species Mystery Supplement
- Dunning-Kruger Effect
- Flowering Time
Category Archives: Update +
Van Reenens Crest
Photos from a recent field trip to the farm Van Reenes Crest (half way between Swellendam & Heidelberg along the N2).
See an earlier article in Haworthia Updates Vol. 7 Chapter 6.
Trevenen & Hesphia Barry
Honey Guide Cabin
P.O. Box 8, Buffeljagsrivier. 6742
084 516 3596

Aloe pumila, Haworthia pumila; what or who is confused?
With some astonishment I learned recently of an opinion that the name pumila as used in Haworthia is confused and therefore should be discarded. There is no doubt that it is difficult to unravel the literature and the usages of the name, but I think there are a number of important questions which should first be answered. Is the name confused or is it us and others, as individuals, who are confused? It is worth asking such questions because names are words in the process of communication and mutual understanding around which all knowledge and its sharing revolves.
If the facts of the matter are properly examined there is a clear path of events. It may be very complex and take many words to explain, but it is there. If a reason has to be sought for confusion I have no doubt that it can be tracked to the door of the International Code of Botanical Nomenclature. My reasons for saying so should become clear.
Paul Forster, who is himself a professional and highly qualified taxonomist, commented on my treatment of the name H. pumila as follows “We do not live in a vacuum”. Here he suggested that I had failed to consult authority and that there was professional opinion available that would have resolved my confusion.
If one thinks about Paul Foster’s statement it is quite evident that:
1. he does not consider himself able to resolve the problem (this is a valid point because to do so does require knowledge of the elements involved, and also requires insights into the intricate mechanisms of the Code which are beyond most).
2. he simply believes that there is someone in the hierarchy who is competent and able to do so.
3. I had not thought there was.
The latter point is simply not true as I have consulted almost every single botanist and non-botanist (yes, many non-botanists are free to dabble in plant taxonomy because of the non-scientific nature of the activity) available to me. A list of names and also authors and published references would fill a few pages.
Turning to the name “pumila” itself, it can be shown very clearly (botanical terminology to mean something quite the opposite, but which I use here in the true sense of understandable) that Linnaeus used the name for four “varieties” of small Aloe. Confusion certainly followed about which of his names should be used, but there should be no doubt about which four these were.
The problem arises out of the Code and how, relating to names for those four “species”, it is now interpreted and by whom. It becomes a juristic problem requiring a great deal of fancy intellectual footwork, complicated by an approach to the code which largely ignores the basic intention to bring stability and uniformity to plant names. It also involves use of words like “validity”, “legitimacy”, “intention”, “relevance”, “strict interpretation” and “according to article…” with gay abandon.
Without going into the very extensive historical detail which includes the fact that the names margaritifera and pumila were used for the same single thing, it is almost sufficient to say that Col. C.L. Scott took the plunge. Defying the predictable response, he took the assistance of Dr L.E. Codd, who was then and still remains a highly respected taxonomists. Together they concluded that because of what and despite what, anyone had done with those two names, the name “pumila” was valid and correct for the species represented by the illustration t10 of Commelin 1701.
They argued that despite the fact that the name had already been used in Haworthia for another species, this did not preclude its correct use for the one represented by the Commelin illustration. The fact that confusion may have arisen and continues to this day, is to my mind correctly laid at the door of Dr W.T. Stearn, paragon and patron saint of plant taxonomy. He wrote a paper in 1938 in which he annotated the names of Salm-Dyck and reconciled them with Berger’s revision of Aloe in 1908. What he did was also to typify the name Haworthia herbacea (Miller) Stearn. In my opinion, had he known anything about the actual species involved, he would have recognised the problem arising from the inclusion of “pumila” in the history of that name and resolved it accordingly. He did not.
After Scott (and Dr Codd) had reached a decision in 1978, I still remained in some doubt using the name “pumila” until forced into a decision when I undertook to write a revision of the genus myself. I had been asked to write a synopsis of the species of Haworthia occurring in the Cape Floral region. This request was by the taxonomists Dr. J. Manning and Dr. P. Goldblatt and I in turn asked them (as had become my routine practise with taxonomists) to clarify the use of the name “pumila”. They contacted Dr Fred Barrie who I presume is a figure in the hierarchy alluded to by Paul Foster, and the following was his reply. “According to the Linnaean Typification Project database, Aloe pumila var. pumila was lectotypified by Wijnands (Taxon 34:310, 1985) on Commelin, Hort.Med.Amstel.2:t.10, 1701. Linnaeus cited this figure under A. pumila var. margaritifera in Species Plantarum (p322) as “Comm.hort.2,p.19,t.10.” Consequently, var. margaritifera and the autonym are synonymous. Var. pumila, as the autonym, has priority.”
There is more to the reply, but it does not address the problem of the different usages of the name “pumila” by various authors until its inclusion in Haworthia by Duval for a species based on a different type.
There is no confusion in this. It is simply a question of how the Code directs that the name should be treated. I was confused over the issue. I did wade through the detail of synonymy with Mr Larry Leach and subsequently with Dr Peter Bruyns. It was evident from this arduous process that, was “pumila” not available for the species in question, the name “maxima” should be used. There is absolutely no reason for confusion about which species I am referring to, nor about the names available or the way in which they have been used since Scott 1978. But I decided to treat the name “pumila” according to Dr Barrie’s response. This suggested to me that Dr Codd and Col Scott were correct and that “pumila” as used by Aiton and Duval was incorrect and did not preclude the use of the name as typified by Wijnands. The essence is that Linnaeus used the name “pumila” as a prime name in Haworthia and it should sensibly remain there.
What is confusing and what should confuse everyone, is that there are persons who feel the need to contest the issue. The need only arises from personal feelings and the fact that the Code has generated this vast arena for endless vain debate. Where a name has so convincingly and obviously been used in the literature of the time, it obviously meets the need. Changing it, or attempting to change it, generates confusion. We do not have to feed on it.
Haworthia limifolia – a conundrum or a lesson?
I was contemplating the problem I see in the way we approach classification, contending that there is no resolution to the endless addition of new names and arguments about their validity and usefulness unless we reach some agreement on method and purpose. The confusion about names, descriptions and identifications that had arisen in 1947 came about for four reasons. Small samples and inadequate exploration, too many unqualified experts, lack of insight into what species actually may be, poor methodology.
There is no excuse for this situation to continue. Sampling is at a very high level and because the other three elements have not been removed, confusion is as great as before. Many names may have been lost from sight, but there is no lack of new ones to replace them. Experts are a problem and their qualifications even more so. In my own experience I have communicated with many highly qualified botanists who have contributed absolutely nothing to the resolution of problems as basic as typification of names, or definition of the word ‘species’. So what then contributes to qualification? This is surely an unbridgeable problem when amateurs have dabbled so freely in plant classification with apparent success. But their success has come from the failure of qualified botanists to put something more substantial in place than a nomenclatural code for binomials and nothing to indicate what those should mean, or who is competent to generate or change them.
Continue readingHaworthia marxii and H. truteriorum in relation to rational classification.
Explanatory note: In a rationalized list of names I wrote and had published in Haworthia Update Vol 7. I made two decisions. On of these was in respect of H. marxii and the other in respect of H. truteriorum. In the case of H. marxi I included it under H. emelyae. In the case of H. truteriorum, I placed this under H. bayeri. In neither case did I have good evidence for doing so, much other than my conviction that a classification is intended to reflect origins and relationships. In formalizing names one perforce is pressured into making decisions that you are not informed enough to make. My main defense is that the authors of the two species were not adequately informed either. I think their account of H. mutica (their H. groenewaldii) at Buffeljags demonstrates this. H. marxii presents particular problems that I simply do not have any substantial data that I can process. In the case of H. truteriorum I did have some to which I can now add. This suggests as Breuer and Marx indirectly indicate, that H. truteriorum relates to my concept of H. mirabilis. I concede that I may be quite wrong in attributing it to H. bayeri. I find it very difficult to see the decision to describe it as a distinct species as a logical scientific action. This article does not extol any imagined virtues or skills of mine. It is only intended to further project my opinion that we urgently need to work towards a classification that does satisfy scientific principle and not novelty or commercial ends. It is also a counter to some very negative opinion aimed in my direction.
At a level above classification lies respect for people and their feelings. Therefore this writing should not be seen as anything but a commentary on the classification process and not how this can also denigrate people as much as honor them. H. marxii is described by Sean Geldenhuys in ALOE 44.1:5, 2007 from apparently 2 populations in a confined area in the north and east of the Little Karoo. Placing it with H. emelyae simply reflects doubts that need to be expressed about how this oddity has come about, and respect lost for reasons not necessary to explain. H. truteriorum is described by Ingo Breuer and Gerhard Marx in ALOE 48.3:54, 2012 and refers to a single population of plants southeast of Oudtshoorn. That this latter population is singular and extremely interesting goes without question except for how it is explained. Despite much correspondence with both authors and after reading their published works, I am not aware that either has a concept of what a species is any different than what may have been held by either K. Von Poellnitz or G.G. Smith. Ingo Breuer particularly has published what he refers to as a species concept of Haworthia. This is nothing more than a long list of Latin binomials and we have to assume that this is then also a list of real species whatever they might be. Gerhard Marx maintains that my attempt at a species definition is so broad as to be meaningless. He has not supplied an alternative definition and I am left with the impression that he makes the same assumptions that “character” differences equate species as does the ordinary uninformed mortal.
H. truteriorum is described in a popular journal (Aloe) that has been approached by at least one botanist not to publish new species as there is no official review process. However, the response was that the journal does have its own in-house authorities that cover the possibilities of scientific lapse. I note that the article is specifically foot-noted to indicate an editorial review and I perforce do not see this as a check on the scientific content. There are some lapses that I will deal with but recognizing that these might not be the same ones that a properly qualified botanist (a minimum of a recognized 4-year degree course), nor those that an experienced and knowledgeable botanical taxonomist may have corrected. More important though is the population itself and where it fits into the overall Haworthia picture.
The most substantial gain that we can make is to arrive at a species definition. It is evident to me that species are complex systems in which there are variations that have arisen from to earth differences and must continue to exist to facilitate response. If there is evolution and thus adaptation and selection, then there has to be something to work with. The phylogenetic idiom was “specialization is precursor to extinction”. Geological and habitat diversity result in plant diversity. This diversity is necessary for survival as the habitat changes. Therefore a species will have a geographic distribution across which individuals and populations will vary and be different from one another. Geology and geomorphology will be strong factors influencing plants like Haworthia that are associated with skeletal soils and rocky habitats. This means that we have to look for associations in respect of distributions and the driving forces that affect even vegetation. Thus in the case of H. marxii (known from 2 populations in a small area south of Laingsburg) the vegetative appearance of the plants cannot be seen in any other species geographically closer than H. emelyae, which is quite a long distance away southwards. The presence of H. pumila in the same area as H. marxii suggests that the Haworthia presence there could extend from the Worcester/Robertson Karoo. Hence H. mirabilis, also present in the Montagu area, cannot be ruled out in seeking a relation to H. marxii. There is much more to this issue of H. mirabilis in respect of its variability and its distribution into the Little Karoo that impacts directly on this suggestion. It also impacts on the real identity of H. truteriorum.
Where I see a real problem with H. marxii is in floral morphology. I gather that the flower very much resembles that of H. marumiana dimorpha. In the same way that the flower of H. pulchella globifera is identical to that of H. cymbiformis incurvula at Plutos Vale, there is a massive problem in drawing conclusions from flowers as a character for the level at which all role players are trying to identify species. But so-much for H. marxii and I do not seriously question it.
The case of H. truteriorum is more manageable. I have been speculating for a long time that H. mirabilis and H. emelaye may in fact be the same species. This may be the proper level at which we should be recognizing species as systems. I extend the argument even to say that it is not inconceivable that H retusa and H. mirabilis are one species. This means that I have to bury my long-held objection to the view that “Haworthia is a genus in a state of active evolution”. My objection being that this is an obvious aspect and that in any case all species are faced with the inevitable need for change and adaptation. But this does not weaken my view that species are chaotic fractal systems that vary around a point of attraction according to the stability of their genetic bases and mutating rate. How strong I am on the technicalities of DNA and evolutionary theory may be problematic, but there is no evidence that other authors even contemplate the issues.
Gerhard Marx is a remarkable observer and I have huge respect for his many skills. Breuer is a really competent compiler too. I do not question what they say about the characters of the plants and their observations. What I question is their knowledge and insight into broader botany and the distribution and variation in Haworthia however much more they know than the above average collector.
My prime objection is that the description involves a single localized population. This in itself creates huge doubts in my mind because on this basis, Breuer’s several hundred species is conservative. I will only dwell on three other points. The one is their very trite “Never before have retusoid type Haworthias been found growing in shale in the Little Karoo”. The second point is the habitat description in relation to geology. The third is the flower and flowering time. The fourth is about the illustrations and the art work.
Haworthia mirabilis was recorded in shale at Barrydale by Smith prior to 1947. It was recorded at two places in shale or shale derived soils prior to 1999 and I can add that I have seen it at two new locations in shale in the Montagu area since. The second point is the description where it is referred to H. bayeri and H. emelyae occurring in quartzite and quartzite conglomerates. As in the description of H. groenewaldii, this is simply a very crude and inaccurate account of a very important issue. I am no geologist but I do know what quartz is and that it occurs in both shale and sandstone formations. Furthermore, I do know that the Oudtshoorn area has an incredible geology with ancient and recent geological formations adjoining as a consequence of faulting and folding. Quartz is Silicon Oxide and is apparently soluble in water at high temperature and pressure formation so that it can accumulate in fissures and bands in parent rock. In both sandstone and shale the quartz varies in purity, and the crystals in size. South of the Langeberg the shales are covered in an extensive layer of tertiary gravels that are far less extensive north of the mountains. But the main point is that the vast quartz patches of the Little Karoo are actually associated with quartz existent in the Bokkeveld shale. The “species” need to be properly looked at in their relation to that complex geology that exists there. Marx and Breuer mention the differences north and south of the Outeniqua Mountains. But the Outeniqua Mountains are just an eastern extension of the Langeberg from the Gouritz River gorge. The geology north of that area is quite different from that west of the Gouritz. This suggests an ignorance of geography added to that displayed for the geology.
Checking my own knowledge and experience of the habitats of the species involved (viz. bayeri and emelyae – with the name picta an anomalous insertion), it is very clear that the statement “quartzite and quartzite conglomerates” is erroneous. There are four geological formations involved and these are the Table Mountain Sandstones, Bokkeveld Shales, and then Enon and Tertiary deposits. In the Heimersriver where the H. bayeri, H. emelyae and “H. truteriorum” are reported, lacks Enon presences. The report of the plants in unfragmented, unweathered upright shale (Bokkeveld) needs to note that the quartz patches in the area are the result of fragmented and weathered shale.
Marx and Breuer perhaps should also take account of H. outeniquensis not a great deal further south in an area that I believe is still unexplored. There is also the mystery of a plant found by Avril Schein in that close area that remains unexplained.
We know from the H. retusa /H. mirabilis interaction that flowering time is no barrier to hybridization and that flowering time may not be indicative of a completed speciation process. Ignoring the fact that we can only guess, on the basis of the scientific paradigm, that we are interpreting and trying to understand an evolutionary process. Breuer has attempted to jump this issue by the recognition of “aggregates” and the two authors use the fob of “the mirabilis/maraisii/magnifica complex. This complex I presume is explained as a list of names only. This I do think emanates from a mis- understanding of a real knowledge of field botany generally and of Haworthia in particular. I say this with great emphasis and conviction because it is something I am still working towards. I have just completed a very thorough look at a the flowers of a very small fragment of populations driven by the expressed opinions of both these authors in diverse places, that flowers are significant with the import that I have ignored them. The fact is that if the flowers are considered then we have a bigger problem than before – not a solution. The flowering time of H. truteriorum that Breuer and Marx cite is very possibly an indicator of behavior rather than a species differentiator. Why they emphasize it is most probably, as J Manning pointed out, most observers (taxonomists) have a subconscious belief that species are things that do not interbreed and so flowering time is seen as such a great barrier to interbreeding that it MUST be a major species indicator. My observation is that this is not true and that species are inherently highly variable systems with great capacity to respond to environmental drama that may threaten their continued existence. The nature of the problem is very well illustrated in the case of H. retusa and H.mirabilis where I contend that a fully developed view of the genus may require that they be seen as one species. Thus the importance of flower and flowering time is part of the myth of a non-existent species definition and is confounded by the concept Breuer and Marx have of the nature of Haworthia species.
There is a very good description of the flower and its character, but the illustration and the art work is weak. The photograph of the flower is of a single dissected flower to show an internal structure that could be of any species in the subgenus and is hardly helpful. That flower does not look to me like the perhaps longer narrower flower of H. mirabilis (real) which is largely recognizable on the arrangement of the petal tips in the bud-stage. This ‘mirabilis’ character should be apparent in the way the petal tips display and we have only a painting to judge this by. Despite Marx’s craftsmanship I am not sure if his flower picture is a true image. I do have an observation on with his outstanding art. Many years ago, photography was a bit of a handicap when it came to illustration of floral and other plant detail. Historically artists were used to capture detail that could not be described or otherwise illustrated. In the present age this is a bit of a myth and it lives on simply because of the skill that is involved in the production of a good piece of botanical art. Marx’s art is up there with the very best. But good art does not equate to good science? Or does it? Botanical art is perhaps not the same as pure art and the intent of either may be quite different. With pure art one could surely be trying to achieve the same goal as science and there must be some trick to understanding the similarity.
My conclusion is to plead for more rational classification and better attention to those things that matter. The fragmentation of a genus by Latin binomials just because of collector and novelty interests, aggravated by commercial implications however slight, is a disservice to all of us.
Very brief note re Haworthia nortieri flowers.
Some criticism about my supposedly having ignored the flowers in Haworthia comes at a very inopportune time. I set aside flowers for the reasons very obvious from the historical record but also because of the considerable problems of similarity in the appearance of the flowers in apparently quite different species. My priority was a geographic overview and a rational basis on which discussion and decision making could be based. It I just grossly unfortunate that other writers and critics seem to be wedded to a classification paradigm locked into the approach that prevailed 70 years and more ago. This in the total absence of a species definition other than the vague acceptance of a zoological one based on interbreeding capabilities. This ignoring the ease of hybridization among Haworthia variants in general.
While I have written an account of flower appearances in a small selection of populations, I also came across these few images I have of flowers in what I regard as the species H. nortieri. I have also added images of a single flower of H. maculata from a population high in the mountains at Worcester that could be seen as a southern extension of the H. nortieri set of populations. Note must be taken of my early contention that H. nortieri and H. globosiflora were the same species, based on my observation of the intermediate appearance of the flowers of a Vanrhyns Pass population. The H. maculata bud is typical of the species in the Southern Cape, whereas H. nortieri has rounded bud-tips.
The flower of the Trawal plant are dramatically different from that of, say, Sneeuberg. It is very understandable that differences like this lie at the base of all the argumentation and confusion that so despoils the naming and identification of Haworthia. A classification has been needed against which to explore and examine these differences. It seems to me totally unnecessary to try and construct another hierarchy of solely Latin names while so little is still unknown.

6158 H. nortieri. Sneeuberg, N Ceres. 

6505 H. nortieri. N Dwarsrivier, Cedarberg. 

6506 H. nortieri. Dwarsrivier, Cedarberg. 

7579 H. nortieri. Trawal Bridge. 
EvJ19550 H. nortieri. Blinkberg.

EA1442 H. nortieri, Middelpos, Bitterfontein

JDV95 ex EA H. nortieri, Kareeberg, Nuwerus.

PVB6146.7 H. nortieri. Komkans. 


6815 H. maculata. Audensberg. 
The reality of Drosanthemum micans L.
Donald A. Levin (2000) quotes Raven, Berlin and Breedlove (1971), who wrote…“our system of names appears to achieve a reality which it does not in fact possess”. I find this a curious quote, as Levin was discussing species concepts and we could ask if species themselves have any reality. There is a lack of a universal definition for the word “species” and I find the recurring reference to “concept” as related to the word, very confusing. Why should the species be a concept, subject to individual interpretation? This is of course if they have no reality and we each create our own. How useful is this for science? Donald goes on to generate his own “species concept” in which he proposes…“that each species has a unique way of living in and relating to the environment and has a unique genetic system…”, and he refers to this as the eco-genetic species concept. (But still we do not know what he means by “species”.) He says that the ecological properties of such species are not uniform within the species and thus not equivalent to the taxonomic properties of species, “which are chosen because they are conservative and stable attributes”. It would be interesting to know just what he means by “taxonomic properties” and I presume he means primarily morphological characters. Certainly there must be taxonomists who utilized or have utilized ecological facets to decide on their species. It must also be recognized that the taxonomist is confined to the material at his disposal for examination and decision making.
My view, to the degree that I can understand Levins’ arguments, is that he has not truly stated the case. The problem concerning the reality of Latin names, is that they have largely been generated by taxonomists who may not have recognized that morphological (“conservative and stable attributes”) properties of a species may vary just as substantially as those of eco-genetic species; and also that these “realms” for each species may be different. From my long experience in taxonomy and the usage of names in communication, it is evident that it is the taxonomic system and taxonomists, which have induced the majority of people who use Latin names in any way, to believe that they do have a reality. The system has been based on the view that Levins apparently has, that a taxonomist determines species by characters or character sets which can be quantified and easily (sic!) used. Perhaps also, that the taxonomist has had enough material to make a decision that is universally true. In the absence of a universal definition it is self-evident that taxonomists and persons, who use the names they give and are given, may have quite divergent views on what those names actually mean and what reality they have. This is especially true if sight is lost of the fact that the characters taxonomist use and have used either morphological or ecological, are simply not as “conservative or stable” as believed. Neither may they be adequately circumscribable or quantifiable. The material examined may simply be inadequate to convey the varied characteristics of the species as it occurs throughout its distribution range.
I would like to use an example to illustrate the relationship of names to taxonomic characters and identification. Drosanthemum micans is an old latin name that was given by Linnaeus for a mesembryanthemum species. The name is apparently based on a Dillenius illustration (fig.1). While the name has been commonly used in the ranks of botanists and horticulturists of my personal acquaintance, there is some doubt that the connection of original illustration and name, to the plants identified as such in recent times may be right. In this case, my use of the name stems from the usage of my predecessor at the Karoo Botanic garden, F.J.Stayner, and he probably obtained the name from identification by Mrs. L. Bolus. I many cases names may come into general use through less authoritative channels. At this point in time the true typification of this name and its correct application is under review, so I will use the name as I know it for a species that to my knowledge occurs both within the Karoo Botanic Garden Reserve, a short distance to the east and also in the developed suburb to the northwest of Worcester, Brandwacht. The name “micans” means “glittering”, and indeed this is the case for this plant (fig.2). The species is characterized by inner and outer rings of bright colour, the calyx bears enlarged bladder cells and the leaves have a grey-blue colour in the summer months. The flowers have outer black staminodes and there is a series of inner smaller petals, often uniformly one-colour, which accentuate the coloured ring effect. There is another species with these black stamindodes viz, D. speciosum, which can be separated from D. micans primarily by the appearance and colour of the leaves. In the former the leaves tend to be slightly more globose and have a greenish-yellow colour. In the latter the leaves are grey-green and tend to be elongated with an uncinate (hooked) end. The black staminodes seem to be unique in the mesembryanthema and who knows if they serve any particular function.
In more recent times, Mrs. Bolus described several other species. One is Drosanthemum bellum that is said to have been collected “near Ceres”, and D. hallii from hills east of Rawsonville. D. bellum was described as pink with black staminodes, and D. hallii as yellow with black staminodes. During a period I which I was interested in these very colourful plants, I came across a population of plants at one spot on the hills “east of Rawsonville” (Die Nekkies, north of the Brandvlei Dam) that demonstrated an array of colour forms. Among these were pink (bellum – fig.3), white, and purple. At the same time I found a population of plants nearby in which all the flowers were bright yellow (hallii – fig.4). More recently I have had occasion to examine this more closely and find many more populations, which indicate the problem that taxonomists have apparently yet to come to grips with if botanists and scientists in other disciplines are to find any reality in among latin binomials. This is that these characters are not diagnostic for the “species”
On revisiting the “bellum” population I found the same array of colours. There were plants with very pale yellow flowers, pink, pale rose-red, white and white shade with purple (figs.5, 6, 7 & 8). There were no bright yellow flowers despite the fact that such bright yellow flowers characterized plants from every other population along the length of the Nekkies. (Here is where Levins could apply his ecogenetic concept!). What was most dramatic was finding some plants with flowers of the micans type (fig.9 in one population of hallii. I observed colour variants in other populations of hallii further to the south-east where the plants were primarily with bright yellow flowers. On revisiting the D. micans population just east of the Karoo Garden, I found that there were plants with the plain bright yellow flowers of hallii as well as pale rose-red to red flowered plants (figs.10, 11, 12).
The question now arises…”What else?” My own approach to taxonomy and plant identification, is that one has to consider not just the ecological associations of plants, which are mostly more difficult to describe accurately than any morphological property, but geographical distribution. Plant species are just like any other material phenomena. They are distributed in space, and they change with time. This is the determinant of their reality. Taxonomy and the decisions that are made are dependent on the material available to taxonomists and it is often simply not substantial enough to establish a practical reality in the application of their names. Looking further afield, I have three more populations of plants from the southern Worcester area namely at Jonaskop (figs13, 14), Lemoenpoort (figs 15,16) and Droogerivierberg (figs.17, 18,19, 20, 21) which fit the micans/hallii mould in terms of all but colour. The population of D. micans on the Droogerivierberg south of Worcester I thought to be fairly consistent in flower colour. It proved not to be and we found a wide range of colours, which extended to white and also included the more typical bi-colour thought to be the real thing. Curiously Mrs. Bolus also described D. leptum from Stormsvlei Pass as white with black staminodes. I have not been able to find it again, but it does seem, in the light of the distribution and variation of D. micans as I have discussed it here, that it is in fact also this species.
Outside of the Worcester area, there is a population north of the Langeberg mountains in the Keerom Dam area, which is characteristically micans (fig.22 – not that single images can represent the variation in the population). Still further afield is a population north-east of Montagu which suggests affinity with the populations of southern Worcester (fig.23).
Then there is Drosanthemum aureorubrum described from near Drew west of Swellendam. It has the characters of micans except in respect of colour. It is inclined to have flowers rather bronze-red-bronze in colour (fig.24) apparently without much local variation, and it should be noted that this variability can only be assessed in the field or by observation of plants mass propagated and cultivated. I can relate that population to three others, one from the middle Breede River Valley at Napky (fig.25), another from northwest of the Potberg at Potteberg Farm (fig.26) and the third from near the mouth of the Breede River at Infanta (fig.27). All well and good, except that the colour does not agree either with D. micans or D. hallii. Nearly everything else does.
The nail in the coffin of a narrow “character” driven taxonomy is a population northeast of the Bromberg Mountain at Stormsvlei in the Swellendam district. I had found these plants when without flower. Visiting the site in September/October when in flower was quite a revelation. There was a range of colours which included the bronze-red of Drew and the southern Worcester and Breede River populations (figs 28, 29). Very significant were a few plants with flowers coloured precisely the same as micans (fig.30). There were also plants with pure yellow flowers, but the yellow was distinctly on the golden-yellow side. I regret leaving out another dramatically varying population eat of Barrydale.
There is yet another “species” viz. D. lavisii, named by Mrs L. Bolus after Bishop Lavis who collected the original specimens ostensibly from between Struisbaai and Bredasdorp. Although Acocks also deposited a collection, purported to be this species, from Northumberland Point, I have looked for it there in vain. I have found three populations of what must be this species with its red flowers, from Napier (fig.31) from Swartjeskop (fig.32) northwest of Bredasdorp and from Soutkloof (fig.33) still further northwest. At first sight this did seem to be distinctive, but in summation I would opt for the geogenetic option. Looking at all the collections and attempting to relate them to the distribution of other plant species (AND their variants), I suggest that D. lavisii is another name for D. micans. I have seen a red-flowered Drosanthemum in the Bontebok Park at Swellendam (fig.34) and Van Jaarsveld and Pienaar report D. lavisii from the Goukou River in the Riversdale district (fig. 35), and these may be related to what Mrs. Bolus described as D. edwardsii from near George (figs. 36, 37, 38) If one considers the geographic aspect and notes the relationship of the distribution of D. micans and D. speciosum then one must perforce suspect that D. edwardsii is the eastern extension of D. micans. D. micans and D. speciosum do grow in very close association, although very seldom actually sharing habitat as they do on Jonaskop. My observation would be that the distribution of D. micans extends into the Swellendam and southern areas, whereas the distribution of D. speciosum is more karoid and it can be found eastward to Oudsthoorn. An apparently vicariant population near Uniondale has flowers which are golden yellow and with long pedicels (fig.39).
Thus we come to the point where we can ask what these names now mean. An aspect of exploration has been interaction with landowners and other members of the public. In this interaction use is made of names. Botanists (non-taxonomists) who have had reason to explore and report the plant species of the Overberg, have used the names lavisii and speciosum. They seemed not to have the capacity to assess if these identifications were right or wrong, and this is not because they were incompetent. It is essentially because from necessity and habituation they apply a species concept which has no reality. When one tries to convey to a lay-person what one is looking for, names clearly have either no meaning at all, or else are linked tightly to an image and association that the person may have derived from book or contact with someone else. It was most notable that in the Worcester/Robertson area, any red flowered mesem was linked to what I know as Drosanthemum speciosum and people found it difficult to conceive that there was a second species also with red-flowers. It proved well-nigh impossible to convey and communicate that either species could have different coloured flowers or that such variants were not different species. Fig. 40 demonstrate some of the variants in D. speciosum, also at Doogerivierberg, but while D. micans occurred in closer association with Witteberg Sandstanes, D. speciosum was on Dwyka Tillite.
A closing point is to refer again to Levis, where he is copied by Charles Craib in the magnificent work on “The Grass Aloes of the South African Veld”. Levins writes…”the eco-genetic species concept has utilitarian value, which is important from an operational concept.” The implication is that taxonomists have propounded a system of names which has little operational value. What then is the sense of taxonomy? Certainly this is what Craib conveys where he propounds a choice of a classification that serves the purpose of the user because the officially accepted system does not (meet the need of an informed user). The fact is that no system will have operational value while there is no universal definition in place, and common realization that a binomial system and names will never have any worthwhile reality unless there is a commonality about what names mean. Taxonomists have to start, with self-examination, to teach the user that names are for communication and have a meaning outside of the very narrow confines of obvious morphological difference, guess work based on limited material and knowledge, or idiosyncratic opinion. While it may be argued that the confused names I have quoted are directly related to the absence of a taxonomic revision of Drosanthemum since Mrs. Bolus last described her species, I use this example to reflect my long general experience with plant names and what people make of them.
Acknowledgment: I must acknowledge the association with Dr H.E.K. Hartmann with whom I have been privileged to communicate over many years, and whose contribution to the knowledge and classification of the Mesembryanthemaceae parallels and possibly exceeds that of Mrs. L. Bolus. Also I would really like to express my appreciation to the many landowners who have taken us on trust and so kindly allowed us access to their property and plants. To list them all would occupy more space than this whole article but I must particularly acknowledge Messrs Poffie and Hettie Conradie of Worcester (Droogerivierberg), Messrs Jon and Cindy Webber of Cape Town (Klippoort, Stormsvlei) and Neil and Saartjie Neethling of Swellendam (Potteberg Farm). Jan and AnneLise Vlok very kindly sent me pictures from Riversdale and Mossel Bay. I do not know how to acknowledge Conservation authorities in terms of their permit system and how these are issued and monitored, nor the paranoid response one gets to the term “collecting”. There is in my opinion a total failure to make any distinction between collecting for knowledge gain and satisfaction of natural curiosity, intelligent plucking for financial gain, or crass exploitation that could lead to degradation of the environment, nor to balance this against doubtful control of development and concomitant destruction of natural vegetation by other agencies. A permit applicant seems to be regarded as a confessed reprobate with no sense or sagacity and an immediate threat to biological diversity. At least this is my impression of how I have been seen. Who wants to draw attention to themselves in this way? Steve Hammer kindly commented on the manuscript.
Reference: Levin, Donald, A. 2000. The Origin, Expansion, and Demise of Plant Species. Oxford University Press..

Fig. 01. The possible type of Drosanthemum micans. 
Fig. 02. MBB7415.2 D. micans, Karoo Botanic Garden 
Fig. 03. MBB7436 D. ‘bellum’, Nekkies Resort 
Fig. 04. MBB7435 D. ‘hallii’, W Nekkies 
Fig. 05 MBB7436 D. micans, Nekkies Resort 
Fig. 06 MBB7436 D. micans, Nekkies Resort 
Fig. 07 MBB7436 D. micans, Nekkies Resort 
Fig. 08 MBB7436 D. micans, Nekkies Resort 
Fig. 09. MBB7435 D. ‘hallii’, W Nekkies 
Fig. 10. D. micans. Panorama. 
Fig. 11. D. micans. Panorama. 
Fig. 12. D. micans. Panorama. 
Fig. 12a. D. micans. Panorama. 
Fig. 12b. D. micans. Panorama. 
Fig. 13. 7433 D. micans. Jonaskop. 
Fig. 14. 7433 D. micans. Jonaskop. 
Fig. 15. MBB7267. D. micans, E Lemoenpoort 
Fig. 16. MBB7267. D. micans, E Lemoenpoort 
Fig. 17. 7472 D. micans. Droogerivierberg. 
Fig. 18. 7472 D. micans. Droogerivierberg. 
Fig. 19. 7472 D. micans. Droogerivierberg. 
Fig. 20. 7472 D. micans. Droogerivierberg. 
Fig. 21. 7472 D. micans. Droogerivierberg. 
Fig. 21a. 7472 D. micans. Droogerivierberg. 
Fig. 22a. MBBsn D. micans, Keerom 
Fig. 22b. MBBsn D. micans, Keerom 
Fig. 23a. D. micans, Gadsfontein 
Fig. 23b. D. micans, Gadsfontein 
Fig. 24a. D. Micans, Sanddrif 
Fig. 24b. D. Micans, Sanddrif 
ig. 25a. MBB7451 D. micans. Middeldrift, Napky 
Fig. 25b. MBB7451 D. micans. Middeldrift, Napky 
Fig. 26a. MBB7498 D. micans, Potteberg Farm 
Fig. 26b. MBB7498 D. micans, Potteberg Farm 
Fig. 26c. MBB7498 D. micans, Potteberg Farm 
Fig. 27a. MBB7545 D. micans, Bar Harbour, Infanta 
Fig. 27b. MBB7545 D. micans, Bar Harbour, Infanta 
Fig. 28a. MBB7493 D. micans, Klipport, S Bromberg 
Fig. 28b. MBB7493 D. micans, Klipport, S Bromberg 
Fig. 29a. MBB7493 D. micans, Klipport, S Bromberg 
Fig. 29b. MBB7493 D. micans, Klipport, S Bromberg 
Fig. 30. MBB7493 D. micans, Klipport, S Bromberg 
Fig. 31. MBB7465 D. ‘lavisii’, N Napier 
Fig. 32a. MBB sn. D. ‘lavisii’, Swartjeskop 
Fig. 32b. MBB sn. D. ‘lavisii’, Swartjeskop 
Fig. 33. MBB7940 D. ‘lavisii’, Ouplaas, DeHoop. 
Fig. 34. MBB7824 D. micans, Klipbult, SE Swellendam. 
Fig. 35a. MBB7572 D. ‘lavisii’, Nature Reserve, Riversdale. 
Fig. 35b. MBB7572 D. ‘lavisii’, Nature Reserve, Riversdale. 
Fig. 35c. A. Vloks D. ‘lavisii’, Nature Reserve, Riversdale. 
Fig. 36. MBB7787 D. ‘edwardsii’, Pinnacle Point, Mossel Bay 
Fig. 36. J. Vlok D. ‘edwardsii’, Pinnacle Point, Mossel Bay 
Fig. 37. J. Vlok D. ‘edwardsii’, Pinnacle Point, Mossel Bay 
Fig. 39. MBB7547 D. speciosum, Uniondale 
Fig. 40a. MBB7471 D. speciosum, Droogeriviersberg 
Fig. 40b. MBB7471 D. speciosum, Droogeriviersberg 
Fig. 40c. MBB7471 D. speciosum, Droogeriviersberg 
Fig. 40d. MBB7471 D. speciosum, Droogeriviersberg 
Fig. 40e. MBB7471 D. speciosum, Droogeriviersberg 
Fig. 40f. MBB7471 D. speciosum, Droogeriviersberg
New finds in Haworthia.
Previously published Cact. Succ. J. (Los Angeles) 84(1): 41-50

Map Legend – east of Swellendam.
1. JDV84/75 Haworthia retusa ‘turgida’.
2. MBB6666 H. retusa ‘nigra’↔ H. mirabilis.
3. MBB7898 H. retusa ‘nigra’.
4. MBB7899 H. retusa ‘nigra’.
5. MBB7897 H. retusa ‘nigra’.
6. MBB7896 H. retusa ‘nigra’.
7. MBB7871 H. mirabilis.
8. MBB7823 H. mirabilis.
9. MBB7909 H. mirabilis
10. MBB7805 H. mirabilis.
11. MBB7801 H. mutica ‘groenewaldii’.
12. MBB7886-7889 H. mutica ‘groenewaldii’, H. mirabilis, H. minima, H. marginata.
13. MBB7722 H. floribunda ‘major’
1. Haworthia marginata and H. minima
The Robustipedunculares is quite a distinctive group within the currently recognized genus. While the four species in the group are generally quite distinct, there are some remarkable complexities that rival that elsewhere in the genus. Recognizable and obvious hybrids are found between H. marginata, H. pumila and H. minima, but there are instances of whole populations that appear to consist of such in-between forms e.g. H. Xmortonii. I have speculated that perhaps rather than have just hybridized, the species have never ever really truly separated in the supposed evolutionary process. There is a vast body of variants that still link them in intimately as this piece will show.
I long ago observed that a small remnant population of ostensibly H. minima just south of Swellendam flowered in November as opposed to the general rule for the Robustupedunculares as late summer flowering. A vicarious population at Brandrivier north of the Langeberg (H. minima ‘opalina’) also flowers in November and both populations have fairly large and white flowers for the species.
I have recorded the normal bluish-green H. minima within the Bontebok Park at Swellendam as well as a very green variant. But in very recent exploration to the south-east we found an even more divergent group of plants that, while varying among the plants, seemed to be hybrids of H. marginata and H. minima (fig. 2 as a single sample and not representative of all). It was September and there were no signs of old or new flower spikes. Kobus Venter, who was present, remarked that the first plant seen was reminiscent of the plants once present south of Swellendam. The plants were large and in exposed situations even colored brownish as does H. pumila. No flowers were present and their color may have shown if H. pumila could have been directly involved at all, while it is essentially its distribution restricted eastwards from the Robertson Karoo by some 20km that reduce the possibility.
What makes the situation more interesting is that nearby was a population of H. marginata that was flowering and the flowers were also large and very white for the species (figs 3 & 4). Added to the fun was a smaller probable hybrid (fig. 5).
Differences and complexities like this do not really surprise me because it is what I have come to expect in my many wanderings in the field. The problem is that it certainly makes classification and any agreement on a set of names very difficult. I just accept it as a fact that plant species can exhibit greater differences between individuals of the same species than between individuals of different species, ridiculous as it may sound. This is because I perceive species as systems of individuals in populations with a very strong geographic component. To actually make a decision it is frequently necessary to determine just what else is growing in the vicinity in respect not only of the genus in question, but also that of other plants. Even the habitat factors need to be considered.
In the case of the plants pictured with this, the Bontebok Park terrain is mostly tertiary gravels, while the habitat we found the plants in was more recent riverine boulders. It is very curious that in the description of H. groenewaldii, the habitat is implicitly described as Ruens Silcrete. I do not think this is true. It is in the true Ferricrete/Silcrete that we found the next and it seems to be these differences in substrate geology that play a large role in generating variation and consequent controversy.

Fig. 2 MBB7891 cf H. minima, Rotterdam 
Fig. 3 MBB7892 H. marginata, Rotterdam 
Fig. 4 MBB7892 H. marginata, Rotterdam 
Fig. 5 MBB7891 H. minimaXmarginata, Rotterdam
2. Haworthia retusa ‘nigra’
I first allied this element with H. mutica because this is how G.G.Smith referred to his collection from Kransriviermond south of Heidelberg. Since then there have been many new collections from which can be gathered that this population is a hybrid one between H. retusa ‘turgida’ and H. mirabilis. There is another collection north of this at Morning Star that appears to have the same parentage but also including H. floribunda. Then there are populations continuing up the Duiwenhoks and then Klip rivers to northwest of Heidelberg, a population between Heidelberg and Tradouw Pass further west and also a population at Goedverwagting south from there. Apart from the Morning Star population (February/March) these are all September/October flowering. There is a population at the southern entrance to Tradouw Pass of the same ilk that is February/March flowering. Then there is quite a distance between these known populations and a remarkable population at Buffeljags south of Tradouw Pass. It is remarkable for the fact that it is lauded as a new species viz. H. groenewaldii when I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February/March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time.
To explore the realities of the situation we undertook two expeditions, one was to Buffeljags to explore west of H. groenewaldii and the other was to the Tradouw Pass area to explore H. retusa ‘nigra’. The first exploration yielded three populations of ‘groenewaldii’, which convince me that despite its flowering time as for H. marginata above, is simply H. mutica in another guise. I also think far too much is made of superficial and trivial differences that are as much characteristic of the variation in the one original population as they are within the four populations and for H. mutuca in its full sense. I consider it significant that H. mirabilis in its more normal non-retuse and dark green form is present in discrete populations both at and west of Buffeljags.
The second expedition was nearly as fruitful. It showed the Tradouw Pass population to be February/March flowering (see figs 6 & 7), while three new populations we discovered between there and the previous records in the easterly Heidelberg direction were Sept/October flowering. They link up to the populations elsewhere that I assign to H. retusa ‘nigra’. An additional find by Jannie Groenewald, for whom that H. mutica viz. H. groenewaldii, variant is named, also took us to a population of what is clearly H. mirabilis (see map) as I know it in its many disguises in the white kaolinic/bentonite clays of the silcrete/ferricrete inselbergs throughout the low-lying Southern Cape. There is unquestionably an overlap of characters between what I assign to H. mirabilis and H. retusa and I consider inarguable that H. mutica is a reflection of a shared gene pool.

Fig. 6 MBB6666 H. retusa ‘nigra’, Tradouw Pass 
Fig. 7 MBB6666 H. retusa ‘nigra’, Tradouw Pass
What this demonstrates again, as does the Kiewietsvlakte populations between Heidelberg and Riversdale, that H. retusa and H. mirabilis are closely intertwined from east to west. There is an added complexity that H. floribunda is admixed along the northern populations and H. variegata along the southern. The admixture of the two species produces H. retusa ‘turgida’ and H. pygmaea in the east and H. mirabilis, H. mutica and H. retusa ‘turgida’ in the west. This is complicated by the other interactions along the northern and southern areas. In the Potberg area it appears that the genetic material of all five “species” is evident in the populations that I have seen there.
While I would like to explain the situation around H. groenewaldii I. Breuer that I interpret as a variant of H. mutica, this should be left for another occasion as too many images are required to support any argument. As it is, the issue of H. mutica ‘nigra’ occupies 29 pages and 79 illustrations in my book Haworthia Update Vol.2 pt 1:50., where all the above mentioned variants are discussed and illustrated. The naming of Haworthias is highly contentious because the species consist of aggregates of small fairly isolated populations that may differ to large or small degree. The populations are in turn also aggregates of plants that can all be identical from vegetative propagation, similar because of low genetic difference or very different from each other because of large genetic differences. Therefore figures 6-13 simply show just a sample of the variation within these four populations in the Tradouw Pass area. The plants vary quite considerably in size too and the one in Fig 8 is nearly 200mm diameter. I have added to the map locations of the only significant other populations that I know of in the area including H. retusa ‘turgida’ and H. floribunda ‘major’, excluding those within the Bontebok Park viz. H. minima, H. mirabilis and H. marginata.

Fig. 8 H. retusa ‘nigra’, Heuningklip 
Fig. 9 H. retusa ‘nigra’, Heuningklip 
Fig. 10 H. retusa ‘nigra’, Heuningklip 
Fig. 11 H. retusa ‘nigra’, Heuningklip 
Fig. 12 H. retusa ‘nigra’, Heuningklip 
Fig. 13 H. retusa ‘nigra’, Heuningklip

Fig. 14 The typical ‘pressure burst’ of white kaolinic soil from under more solid ferricrete where H. retusa and H. mirabilis are commonly found.
Perhaps I should close by explaining that I have dropped the use of any rank below that of the species name. I simply am suggesting that we recognize the need for a trinomial system without the pretension of status, and more greatly honour the binomial as an entity of a greater significance than we may know. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable specimen that is used to anchor the Latin name.
Acknowledgement. Any proper excursion into Haworthia territory always requires acknowledgement of landowners and I thank Jaap Viljoen and Jannie Groenewald for organizing that and for their company. I was also glad to have Kobus Venter along who had persuaded me to show him some of populations known to me on promise of new exploration.
Note.
Cross seasonal hybrids observed are-
H. retusa turgida X H. floribunda Blackdown, Heidelberg.
And also Platjieskop, Riversdale.
H. pygmaea X H. floribunda Coopert Siding, Albertinia.
H. mirabilis X H. retusa Soetmelksrivier, Riversdale
H. mirabilis X H. variegata Stoffelsriver, Swellendam.
References. I need to record that Harry Mays of Alsterworthia kindly undertook the non-profitable publication of 5 volumes of Updates (2-6) between 2006 and 2009. Vol. 1 was published by Umdaus in 2001. These volumes were the product of research to validate or correct what appeared as a formal revision in Haworthia Revisited, published by Umdaus in 1999. The description of H. groenewaldii appears in Alsterworthia 11.2:15 (2011).
What do collectors need?
Previously published in Alsterworthia International Volume 12, Issue 2. July 2012.
I ask this question because too often the views of the collector are espoused as an excuse or defense for some or other argument about classification. It has often been said to me that collectors are not interested in taxonomy and they are at the most, happy just to have a name. This argument does not impress me because as a society we have a trust and a belief in science and whatever is written, outside of fiction, should seriously address the truth. It should not matter what the reader may want to make of the product other than that the reader may just by chance really want to know and understand something. On reflection, one writes for the reader who must surely be reading because they want to know something, and names are the key to the “something”?
This is why I have responded to reviews of my writing that have been published at various times. I have written as a communication and am glad to know what the reception or rejection has been. Recently Steven Hammer wrote what is listed under the title of “Book Review” comment on a recent book by Ingo Breuer and of Update Vol. 6 by me. It is a wonderful piece of prose and worth every bit of reading and appreciation, but it does not pass as a Review. Or does it? I feel that it has a few mistakes as well as passing over the very real differences between the two publications. So, I wrote a response in the way I treat any publication as an invitation to think and form an opinion; and express it. Passing a draft of this response to a competent observer, I got this reply…” Fortunately, there is little expectation of a review. The point is: was the review positive or negative? Did the reader learn something and gain deeper appreciation, or not? Will they buy the book, or did the review satisfy their curiosity? For most readers, the details are unimportant, as much as you may hate this very concept.”
Why I should hate the concept of most readers regarding the questions of detail unimportant I do not know. But I do think the accuracy, in respect of detail or general, is very important. What my commentator was implying is that the review met the requirements that he was suggesting, and he added that my response was “nit-picking” and would only be seen as criticism of someone who is widely held in high esteem. The fact is that Steven Hammer is also held in very high esteem by me and I am so glad to be able to say that he expressed to me personally that his “review’ was rather a literary fantasy. What Steven does comment on is a view of the needs of collectors. That they care little about schemes of classification and that labels are necessary irritants. I do not question the truth of this view. But would not accept that this is a justification for the imposition of just any kind of scheme because that is what a writer wishes to propound for reasons of his /her own.
These then are the points I made in my response that I think Steven should have addressed. The ‘mistakes’ are … a). The Audensberg population was actually shown to me by Elsie Esterhuizen many years ago and it is not the place where any haworthiophile would ordinarily look for plants. b). The reference to Drosanthemum bellum is odd because Steven describes this as a “niche-sensitive species”. This “species” is at the heart of a very long and detailed story of Drosanthemum micans that I once wrote and lies unpublished. I would surely have used this as an example of the way in which botanical science has also failed us. D. bellum is a pink flowered variant in a much-localized population of D. micans that also has white, purple and red variants. This tiny population sits among a larger widespread population of yellow flowered variants that go by the name of D. hallii. This is turn has variants that include the typical bi-colored flower of the older D. micans that is common north of Worcester. Further variants occur north-east of Montagu, to Oudtshoorn and then south to Mossel Bay (D. edwardsii) back west to Bredasdorp, (D. lavisii, D. aureopurpureum and D. croceum?). The problem here is the failure to establish what is meant by “species”. To refer to D. bellum as a species is a misconstrual of science, or an example of the liberties that are taken with Latin names – botanists and collectors alike. c). Chameleons. Wonderful words of Steven’s, but not quite complete. The story about chameleons’ parallels that of the very low non-tech problem of impossible identification even when there are heaps of “characters” to use. It fortuitously exposes the probability that we are being led up a garden path by high-tech. I have used chameleons in the same way that I studied Oxalis ultimately demonstrating that species are complex systems of variables! d). Kaboega is not the only area I know exceptionally well and it also figures in practically all the other volumes of Updates of which there are five. I doubt if these have ever had much coverage, but they are an account of my voyage of exploration and discovery that is the concern of mine in respect of omission. In the Updates I discuss the populations and their variants as they occur at many different places and show this impact on the application and use of Latin binomials. There is a prevailing misconception that this is only a problem within Haworthia. I show that this is not so. I also make several references to the fact that Haworthia is by no means an integral single genus and that the nature of genera in the Alooideae needs proper attention based on a lot more than the fact that the Haworthia subgenera have small flowers. e) “Shaggy dog” for H. mirabilis Ballyfar is not a name I coined but Steven himself.
I do think the omission is in the comparison of the books where there is in fact none. My Update Volumes revolve around the way that science has let us down to the extent that any pretender can take up the mantle of taxonomic expert. Botany provides no species definition and hence the Latin binomial is not required to carry any meaning other than a guise of authority. Whatever collectors may require has no import whatsoever in a process of classifying plants as biological entities. They are focal points for the collection and storage of knowledge indicated by Latin binomials and these are not simply and only intended as labels. Even I recognized this as a child when I wanted to know what the plant actually was that my father called H. chalwinii, and where it came from. Every collector who refers to names at all surely expects and believes that there is some mystic or real knowledge associated with the names he is given and uses. It is an injustice to any collector to coin Latin names outside of the context of science where the collector is entitled to believe they belong.
The only predicable thing about Breuer’s system, which is a watered down version of a much more focused and detailed one by Hayashii, is that there are going to be a lot more names. This is not only when someone else climbs the Audensberg, or recollects the Sandhills population, or drifts across into the Heatley Peak area. Throughout the Updates, I warn and demonstrate that character fixated taxonomy may be very misleading. Vavilov was a Russian botanist who pointed out that variations in a genus may be expected in all the members of that genus. Species are therefore to be seen as systems that are natural assemblages of plants that can be associated in respect of ALL the forces, factors and features that generate them – not propagules of single clones that fill availability lists and price catalogues. Drosanthemum bellum is just such an example of how Latin binomial names are used to describe variation within species, rather than to properly organize the basic entities that make up the entire living system.
I have, even in the Updates, shown how the watchdogs of science let us down. I have tried to communicate my experience and observations to a wider and expectedly interested audience. This, in the hopes that it would lead to greater understanding and comprehension of the problems of finding names as the backbone of communication, appreciation and understanding. It is a huge disappointment to me that I have achieved very little other than to grow wiser myself. One of my many critics makes a show of taking up middle ground between me and other Haworthia taxonomists. My response is that taking up middle ground between myself and the ignorant is not going to be very productive. In the first place there is not much space there as I am quite aware of my intellectual limitations. In the second place I have not actually been all that certain that my overview is entirely correct. Despite being credited with a lot of field work (and no good sense to go with it) I am extraordinarily aware of how much I have not seen. This adds to my discomfort as I see a proliferation of new names, gaily forgetting the multitude that I moved aside in my Revision. These are often based on propagules from my own collections (concealed by the creation of new collecting numbers that are not mine). I recognize that the only predictive element in this kind of science is that we can expect many more Latin binomials in a collector driven system rather than one of botanical science. So indeed I see no change from the failed methodology of Von Poellnitz, Smith and Scott.
There is no comparison at all between the two books that a true review might have suggested. One book (Breuer) is a collector’s guide to a limited range of variants (albeit 336?), while the other (Bayer) is an account of a very wide range of variants and a hypothesis (not a statement) of how they are related. The latter also throws some light on the universal quandary knowledgeable observers soon come to experience viz. Elton Roberts in the same edition of the Journal where he questions the identity of Mammilaria lasiacantha. The problem he has is a classification fixated on superficial small differences rather than one based on the realities of the variation that should be expected in any system arising from, and actively responding to, differences related to time and space. The essence of science is that an experimental method is applied to a sample and repeated, the result will be the same. If everyone is coming up with a different plant classification, it should occur to us that there is something wrong with the method and perhaps also the hypothesis that is being tested.
I am curious why my commentator dismissed all the above as “nit-picking” when myself I feel that they deal with the most significant elements of writing at all. Especially, they touch on the very core of why we even classify and name plants in the first place. My response should not be seen as a criticism of my commentator, or of Steven, a remarkable man who is also very dear to me. I also respect him enormously for his empathy with plants because if there is anyone who projects my view that this is a conscious creation all the way down to its rocks, it is him. There is surely purpose in creation if only that we should seek and find what that is.
Righting some misconceptions.
The article by Gerhard Marx in Haworthiad 26.2:41 is really excellent. There is however some behind the scenes innuendo as well as repeated use of words like ”ignored” and “neglected” that need attention. There are furthermore comments on “substantially different and unique characters”, “forcing” elements into synonymy and use of words like “truthfully” and “knowledge of the genus” that distorts a reality that is historically under stress. This is totally unnecessary in an article that otherwise contributes so much in the real area of interest and information.
What we first could deal with is this vexing question of what science is and who is a scientist? In the issue of Haworthiad the editor feels it necessary to state that he is not a scientists and both Al Laius and Stirling Baker imply that there are things that the uninformed dare not comment on. Let me make it clear that most of the writing and classification of Haworthia has been done by authors with no formal training in biology whatsoever. I could fill the whole of an issue with detail of how trained and experienced botanists (scientists) have made mistakes in their small contribution to our “knowledge of the genus”, or simply confounded it.
What the Gerhard Marx’s article actually does is ignore history and written knowledge of the genus. He does profile himself on Facebook as a “succulent botanist” but I doubt if this meets the South African Association of Botanists requirements for membership. Many taxonomists have not been scientists at all, and one cannot fault the enormous contribution that most ordinary people (education-wise) have made to botany. So who am I then? With an MSc and a thesis on morphology and taxonomy, am I a scientist? Does training and a formal qualification mean anything at all? Am I really just another writer? Perhaps not. But…
My first bone of contention is the use of the word “ignored”. This is because it is combined with the notion of unknown transitional elements. This is plainly misleading and incorrect. Von Poellnitz described H. mirabilis ‘beukmannii’ as a variety of H. emelyae probably as the first clue to some sort of transition between elements south and north of the Langeberg and Outeniquaberg. Von Poellnitz cites several specimens from north of the Langeberg that he confused with H. pygmaea and what I later recognize as the totality of H. mirabilis. As long ago as 1976 I published a map in Excelsa, replicated in my Updates, to speculate on the transition of the southern retusoids into the Little Karoo as far as Steytlerville. In a recent treatise available to all I explained the remarkable similarity that I perceived in H. pygmaea in the Herbertsdale area to H. emelyae a relatively short distance away north of the Gouritz Gorge. Many years ago and lost in my writing is the extraordinary similarity of H. emelyae ‘major’ and H. mirabilis ‘paradoxa’. While surely there must be a record there too of the extraordinary similarities that I observed of H. emelyae ‘multifolia’ to H. mirabilis ‘sublineata’ as well as to H. mirabilis ‘heidelbergensis’ and even to H. rossouwii.
As an aside I must just draw attention to Al Laius’s comments on what he refers to as H. heidelbergensis and H. magnifica. This is because Marx also uses names like átrofusca’, ‘magnifica’ and ‘mirabilis’ with no appreciation of the realities of these names. This brings me to the second bone of contention – the one of “neglect”. One can only do so much, and if Marx can concede that then he can approach this from a more objective angle. I have been fairly limited by opportunity and finance apart from the enormity of the task, but I can and have pointed to a multitude of situations that are no different from the area in the Little Karoo that Marx has easy access to. I have not felt it necessary to venture there where activities of collectors and others have made it too often to ask the indulgence of landowners. Also, when there is just so much else to do. But one of the situations I must refer to is that of the “species” H. groenewaldii, the description of which was also instigated by Marx. The explanation of this taxonomic deviant occupies almost all of Haworthia Update 7 that is available in hardcopy and also on the internet. This explains some of the aberrations that filter through to Marx’s article.
Does that bring us to the question of “substantially different and unique characters”. My MSc thesis had as the subject the nature and significance of complex morphological structure – albeit in entomology. So I actually do know a little bit about characters. I also know a little bit about the use of DNA strings as character sources. More practically is the work that I have done in respect of characters in Asclepiads, in Drosanthemum, especially in Oxalis and even in chameleons. A scientist unfortunately cannot restrict him- or her-self to what they think are the realities in one small area. There is far more to life than Haworthia and it does not have any special problems. There are any number of substantially differences and unique characters even in individual plants. Marx throws this all away with a rather mixed final sentence and reference at the end of it to a “very consistent set of measurements”. Stephen Gould once said that the strongest statement that one could make in biology was “hardly ever”. A consistent set of measurements in biology is spelled out in the science of statistics viz. biometry. Perhaps that article by Loucka and myself dealing with the statistics of H. mirabilis ‘sublineata’ needs to be unearthed. Some variation in Haworthia mirabilis var. sublineata
So this takes me to the other bones of contention viz. “forcing” “truthfully” and “knowledge of the genus”. I have written elsewhere that the taxonomic system is at fault, and I will not even try to defend that simple statement. It is. One of the difficulties is that things that are neither here nor there have to be unequivocally assigned to a taxon. If this is coercion as Marx implies, then he is unaware of a very self-evident truth. There is no place for the chemical equilibrium equation or the subtle variants that Marx even describes. The sorry fact is that formal botany and the ICBN does not actually provide for “aggregates” and this is just a ploy to evade the issue of the definition of “species” and what it truly means. Manning as a professional botanist needs more than ordinary opinion, in a very small field of interest, on which to base a formal list of names for a national catalogue of 23000 names. To treat each genus in that list on the basis that Breuer and Hayashi have done would be unthinkable.
In closing we have this fudging statement…”Haworthia will remain a taxonomical argument problem for many years to come”. It surely will when writers, editors and readers prefer the mileage they get from disagreement instead of properly informing themselves and working actively towards a common goal with “truth” as a commitment rather than as a word to discount what is already suggested.
Comments on H. marxii
In An honorary Ariocarpus in Africa – Notes and updated information regarding Haworthia marxii (Alsterworthia Intenational Volume 12, Issue 2) Gerhard Marx and S. D. Gildehuys respond to the inclusion of H. marxii under H. emelyae by Manning and Bayer in the Rationalized List of Names.
The reason for my dumping H. marxi was purely political as I explained elsewhere. It is just my opinion that there were and are shortcomings in the author’s whole approach to names in Haworthia. So I expected some attempt at vindicating themselves like this. I have no problems with considering H. marxii as a species. I would simply prefer that it remains an element that desperately requires an explanation.
My problem is with statements like this …”Typical H. archeri can be found only … at Viskuil (JDV89/62)”. I described this species and from where it occurs at Ghaapkop and at Baviaans Station just east of there (and Viskuil is just a few miles further east). I later placed it with H. marumiana because of several other oddities widespread and also because of the very wide distribution and variability of H. marumiana. I have since found it also at Lospersberg. Marx has previously been on at me about the relation of H. archeri to H. marumiana. I did not realize that he was unaware of what actually constituted the knowledge of either species.
There is also mention of a collection of PVB1405 as from 20km SE of Rooiberg Pass. This is a bit of a mystery to me as I thought I had noted this collection of PVB from east of the Floriskraal Dam and was for a long time the only possessor of the single plant in cultivation. This collection is cited in my Revision as are the two “archeri” collections noted above. I am sure I noted this collection 40km ESE Laingsburg. It is a mystery because it is north of the Witteberg and Swartberg and is tied in with another population that Bruyns recorded at Baviaanskloof. Every indication is that it is connected to a population of mine at Scholtzkloof, Prince Albert; and so with things that I pondered may be a marumiana connection i.e. ‘var. viridis, or even connected to monticola or more probably, H. vlokii.
Another statement I must make is about the significance of flowering time, as the evidence is that it is not as meaningful as the authors imply. There are hybrids and population evidently arising from hybrids in these interactions – retusa/mirabilis, mutica/mirabilis, floribunda/pygmaea, floribunda/retusa,and herbacea/reticulata. The latter is not across a seasonal flowering difference but the others are.
The large gap between Montagu and Rooinek Pass is reduced somewhat by the old populations of H. mirabilis northeast of Montagu at Dobbelaarskloof and by the new population recorded near the Pienaarskloof Dam. Also relevant is the distribution of H. pumila. So maybe exploration has simply been inadequate. I was and am fully aware of the distance issue.
Regarding flower morphology. Similarities in the flowers of otherwise very different things, is just as significant as differences in the flowers of things that may in fact be very similar. So these small differences should not be punted without far more attention been given to what actually constitutes difference. The few pictures of the flowers that are provided are hopelessly inadequate to base any opinion on. The ultimate issue is that the authors are using idiom, method and logic that have been in service since before Von Poellnitz, Smith, Scott and the rest of us. It has not worked. It is not going to resolve the complications we can find in situations like between say H. mirabilis and H. retusa, or H. arachnoidea and H. mucronata, or H. cooperi and H. cymbiformis, or H. cooperi and H. bolusii var.blackbeardiana. To name some of the more obvious interacting species or elements if we do not know what species are.
Haworthia maculata ↔ Haworthia pubescens, MBB8002 Cilmor.
In Haworthia Update Vol 9 there is a report of a population MBB7997 identified as Haworthia pubescens from north of the Cilmor wine cellary. This is approximately 2-3km southwest of the type locality for the species. I noted that the plants have less spinuliferous leaf surfaces and there is a degree of surface translucens and maculation (spotting). I also presented 3 pictures of MBB7271 of what I identified as H. maculata from south of the Cilmor cellar. When I first visited this locality I had no problem identifying the few plants I saw as H. maculata on account of their marked spotting. However, on a recent visit we struggled to find plants at all and the few plants we found were too embedded in rock cracks to make any worthwhile identification. So we revisited the site to explore more extensively and located a large number of plants higher up and slightly west of our first sightings. These plants are illustrated here. They incline more to H. maculata than the plants at MB7997 and I have accessioned the population as MBB8002. There is the usual expected large variation in respect of superficial and observable characters. The plants can be proliferous and cluster, more so than at MBB7997. Similarly the leaves can have more translucens and even less spinuliferousness of the surfaces. Some plants have few and quite thick swollen leaves while others may have more and very slender pointed leaves. I have not observed the flowers and really do not expect them to make any difference to the problematic classification of populations that again are neither here nor there in a narrow concept of species. H. herbacea occurs at all four geographic positons at a radius of about 2km. At the brickfield to the northwest as well as just northeast of the Brandvlei Dam wall it is evident to me that there is a transition between H. maculata and H. herbacea. I did report the known distribution of H. maculata in Update 9. While there is no suitable habitat between Die Nekkies hills at the Brandvlei Dam and the Audensburg or Kanetvlei, there is unexplored suitable habitat southwards to Moddergat and Hammansberg. There is no evidence of H. maculata eastwards to where H. reticulata is known about 15km east on Ribbokkop. Westwards no Haworthia is known although G.J. Payne did inform me that he had observed plants in the hills immediately southwest of the Dam at the now submerged hot spring in the Brandvlei prison area.

View 1 From Cilmor to the northeast. 
View 2 From Cilmor eastwards.
The submitted pictures include two views. View 1 is looking north of east across the Breede River to the Sandberg where H. pubescens occurs. Its full occurrence on those low hills is not known and this I will explore soon. View 2 is looking eastwards looking at a Dwyka Tillite hill across the river in the upper right. We found no Haworthia on that hill although both H. pumila and H. herbacea are present on the smaller rise to the right and behind it – also Dwyka. H.herbacea is very abundant on a Dwyka tillite hill about 10km to the south. The corresponding hill on the left is Ribbokkop where H. herbaea, H. reticulata and hybrids are present, and H. arachnoidea also occurs. The limits of H. mirabilis are the higher hills in the background viz. Rooiberg, Gemsbokberg and those are Witteberg sandstones.
Addendum Haworthia pubescens MBB8011, SW Sandberg
Addendum. H. pubescens MBB8011, SW Sandberg.
I need to point out that there is a still earlier article which covers Haworthia maculata (Haworthia maculata <–> Haworthia pubescens) that lays the basis for this discussion. In that article I note the position of the Sandberg to Cilmor and DeWetsberg and intended to include the Sandberg H. pubescens in that article. We could not get landowner contact and so that fell away. However, this problem was overcome and we first explored a Dwyka Tillite outcrop southeast of Sandberg. There is a vast accumulation of windblown sand on the first hill and we saw no Haworthia. There is a smaller hill further to the southeast that is also Dwyka and erosion exceeds wind deposition so smaller non-geophytes do quite well. We found both H. herbacea (see fig.1 MBB8014) and H. pumila there. From there we went to the southernmost point of the Sandberg. A misjudgement landed our vehicle in mud and the drama to get out limited the time we had to explore. We found a lone H. herbacea (fig. 2 MBB8012). Returning a week later we approached the Sandberg from the southwest, and almost immediately on reaching the top we found H. pubescent. Fig. 3 is a view towards Cilmor and DeWetsberg where the plants appear to be intermediate H. pubescens↔H. maculata. The picture is useful to get some idea of the role of geographic and geological considerations. The high mountains in the background are Table Mountain Sandstone and no Haworthia is known there. I am not certain that this is true and G J Payne did tell me that he had seen plants on the extreme lower right and south of the Brandvlei Dam. But also on the absolute distant and absolute left, is the Riviersonderend Mt. That is also TMS. The deep Wolfkloof Valley behind that is the locality for the much unexpected H. herbacea ‘lupula’. (These inverted single commas are not entirely necessary but I use them to underscore my informal use of names that have less reality. The var. lupula is real). The mountains ahead of that last line are Hammansberg on the left and the Moddergat to the right. Between there and DeWetsberg has not so far turned up Haworthia, but this is an exploration problem. Behind the DeWetsberg is also underexplored. H. herbacea does occur between DeWetsberg and behind the mountains on the low right just in the picture and also east of the brickfield out further right. H. maculata is only known in this area along the Nekkies north (further to the right) of the Brandvlei Dam just visible in the picture.

01. MBB8014 H. herbacea, E Sandberg. 
02. MBB8012 H. herbnacea, Klipkranz Camp. 
03 . View west of south from Sandberg.
Prior to this exploration H. pubescens was to me only known from the northern part of the Sandberg that lies south of a road going eastwards to Eilandia between Worcester and Robertson. Here H. herbacea does occur on the lower northwest warm slopes. H. pubescens seems to occur only on the upper two ridges and H. herbacea is not known to intermingle with it. This is all Witteberg Sandstone that as a formation overlies Bokkeveld Shale and underlies TMS.
Coming back to the southwest corner of the Sandberg where we found H. pubescens. The plants seem very similar to the species as it occurs to the north. They were very cryptic and often in shady rock retreats where they were really hard to see. It was mid- to late-morning when we were there and the plants were not going to be better exposed as the sun moved further west. Although there was very suitable well-drained habitat lower down on the shaded east slopes, there were no plants and I speculate that this may be because the plants may need the cooling effect of wind movement up on the ridge. The pictures tell the story of variability in respect of a whole range of leaf and rosette characters.
It is worth noting fig. 43 of the dead remains of a plant under a clump of restioid. It seems that seedling survival is closely coupled to early protection giving rise to the concept of nurse-plants. Plants are often very difficult to find because they are so hidden beneath accompanying vegetation. But they do need light and the dynamics of vegetation growth and densification must have quite a big impact on the ageing and survival of plants. It raises again the question of how long do the plants live? For plants like Aloe ferox and A. dichotoma I do have a real experience of a lower limit of about 35 years and a top limit in the several hundred. In the field, the plants seem comfortably ageless.
The really interesting part is this. While I was busy tediously cleaning a plant to photograph it, Daphne called to me to come and see a lighter green plant she had seen. Moving in that direction I saw a plant that registered as H. herbacea but with some hesitation and doubt (see fig.4). I then went to see what Daphne had observed. They seemed less obviously H. herbacea but that seemed to be a logical and conservative opinion (see figs. 5-11). That was until Daphne found two adjacent rosettes at the foot of a restioid clump that left me in no doubt that they were hybrid H. pubescens/maculate (see figs 9-12). These were in bud whereas H. pubescens plants showed no sign of impending flowering. Note the buds are less well developed than MBB8014 further east, even if possibly insignificant. Going back to the other plants we confirmed my doubts. They were a lighter colour and apparently softer texture that we would have expected in H. herbacea. These were the only plants we saw in a space bridging the occurrences of plants of H. pubescens.

04. MBB8013 H. herbaceaXherbacea, Sandberg.







05-11 MBB8013 H. herbaceaXherbacea, Sandberg



































12-46 MBB8013 H. pubescens, SW Sandberg.
We did not explore the western slopes where habitat would have been more suitable for H. herbacea and I expect it does occur there. What puzzles me is that so frequently have I found very distinctive hybrids between species in close proximity and very seldom where the species are some distance from one another. I cannot say I have ever found a hybrid in the clear absence of both parents. The example of Astroworthia bicarinata at Lemoenkloof, east of Barrydale, may be an exception where only Astroloba corrugata (syn A. muricata, A. aspera) is present but H. pumila apparently not. Hybridization is thought to be an important element in the “evolution” of new species. I doubt this as it is quite evident that separation into two species is a pre-requirement. If new species have evolved in Haworthia by hybridization, how did they evolve as such in the first place? The answer to me lies in the continuities between populations. I observe, and have experienced of expected continuity between populations. While the Cilmor populations are thought to be H. pubescens↔maculata it cannot be said anymore that they are hybrid, or populations where the morph or drift to discrete elements has not reached a conclusion. The latter is more likely. As there is already apparent geographic continuity of H. maculata and H. herbacea, I was expecting some evidence of a similar relationship between H. herbacea and H. herbacea. So here it is. Hybridization as a factor in speciation in Haworthia does not seem to very likely. It confirms for me that there is a fractal “chaotic” order to species in Haworthia and the reality is that a view of many truly discrete species is a fabrication and a very ill-considered view.
Acknowledgement:
We are always greeted with such kindness and helpfulness that we might have expected this from the Sandberg landowners too. It came in no small measure. Driaan Griesel was most enthusiastic and interested and also helped us with extracting our vehicle from the mud on the one occasion, and then jump-starting it after a flat battery on the second. Our imposition did not so much as touch his view of the day.
(ed. – Bruce made another visit to Southwest Sandberg on 9 December 2012 and includes the following flower pictures. He makes this comment; personal correspondence 27 December 2012.)
I am actually not sure at all about flowering time now. I used to be quite sure of being able to collect seed of pubescens mid-Nov. But I observed at Humansdorp that gordoniana peak flowering could be out by 6 weeks. In any case the plants can produce successive spikes so one can get delayed flowering and added to that energy in the first or the second flower set. I know mirabilis at MacGregor can flower from Nov. thru to March while at Montagu mirabilis can flower as late as April/May. Retusa and geraldii are quite happy to produce flowers in either Spring or Summer and Kobus observed that splendens did that too. Maculata can flower from Sept. thru to late Dec. And each population does its own thing.


































































.
Genetic history – the place in Haworthia classification.
Preamble: Maybe the difficulties and strife in Haworthia classification is that we do not know what it is that taxonomists do. Taxonomy is the academic discipline of defining groups of biological organisms on the basis of shared characteristics and giving names to those groups. The scientists that do taxonomy are called taxonomists. The names taxonomists give to species don’t just tell us what they are called, but also tell us about how they are related to one another. In the following article Bruce Bayer shares his views that Haworthia taxonomy should be about more than identification and descriptions, rather the interpretation of the characters that reveal genetic relationships and evolutionary pathways. ~ Lawrence Loucka, 7 September 2012
Genetic history – the place in Haworthia classification.
By M. B. Bayer
This is a huge subject and I cannot by aptitude or knowledge attempt to cover it fully. However, it is so basic to plant classification and Latin naming that we all need to have some background understanding. The entire object of classification boils down to arranging living (and once living things) in a scheme that follows the theory of an evolution from simple life forms to the present array of creatures. This is because it is thought to be the logical way in which knowledge of the said creatures should be organised i.e. according to the way in which they traveled the creative way. All classification is simply a way of organising information so that one can generalise from the singular to the plural.
Whether one believes in such a progressive process or in a spontaneous divine creative event does not matter. All the evidence points to change of some kind and if we call it evolution does not alter anything. It is argued that there is in fact no direct proof that, say, man evolved from a primitive primate. That may well be so, but there are many fossil primates that are speculated to fill that gap. The case is that evolution is a scientific hypothesis and science does not, or cannot claim, that this is proven. A scientific hypothesis simple stands as a possibility or probability of something known. Such a hypothesis can only be proven untrue, and then it is replaced with a new hypothesis that stands as the accepted truth.
Whatever, the ultimate truth of the matter, classification in science is directed at reflecting how the different life forms are related to each other in respect of change? In the acceptance of evolution the process of change is the genetic history of the organisms and is referred to as the phylogeny. It is often depicted as a branching tree with the branches originating low on the trunk as ancient origin, to terminal branching for recent.
My experience with classification comes with education and qualification in entomology (insects). It so happens in insects (and higher animals) that there are many characters and structures that can be used to examine the process of change and modification that might go with evolution (change). The word “homology” is used. So, say, the wing of a bird is likened to the forelegs of a dog – they are said to be homologous structures. The change in structure from that of a segmented worm (eg. Onycophora) to say, a wasp, can be tracked through a whole range of insects. The head of a wasp is seen to be derived from 6 primitive body segments, the thorax from three, and the abdomen from several. There are muscles and nerve endings and sub-segments that are all evidence of these connections. In entomology, there was a stage when a qualification in insect classification required the student to also present a phylogenetic scheme to explain relationships relative to an origin. This all based on morphological characters, their homology and their interpretation as primitive or advanced. What is really odd is that too often a classification comes to deny the variability that a concept of selection and adaptation that evolution rests on, is denied.
In plants there is no luxury of musculature, or nervature or even detailed bone or segmented structure on which to base good homology. Botanists are therefore very hard-pressed to properly derive phylogenies or explain this genetic history. Nevertheless a very serious and honest effort is done to do this. The process of cladistics was derived from entomology (Hennig) and became the standard for the student in botanical classification. Students were required to evaluate characters on a basis of primitive and advanced, and construct cladograms that illustrated a hypothesized process of evolution. The advantages of the method over conventional decision making were that it is said to be an objective way of arriving at a conclusion rather than personal subjective decision making. With the complaints against my use of words and language, it is worth noting the remarks of a leading botanist who said that the methodology of cladistics was just the use of a new language that did nothing more than strain good-will. Agreed.
With the arrival of DNA sequencing, a whole new ball-game was created with each amino-acid base pair representing a character. So DNA sequencing has become the modern standard for the construction of phylogenies and genetic history – the phylograms. So much so that the article in Taxon 56:645-648, 2007 by A.B. van Wyk argues that the phylogram represents the process of phylogeny as the genetic history as opposed to the cladogram that attempts to construct the genetic history. I personally think that this is intellectual wizardry and the literature references in the Taxon article only expose the mystery that surrounds classification. It takes classification wholly out of the reach of any but a polymath. Basic to the whole issue is that scientists still have not reached any certainty about what a species is. The theory of evolution is not disproved. Van Wyk has said
Plant taxonomy, despite all its impressive achievements towards phylogenetic reconstruction, will then risk being denoted as yet another ivory tower science – a pursuit disconnected from the practical concerns and needs of everyday life, esoteric, over-specialized, its classifications of little practical use to the majority of end-users”.
It sounds very impressive, but I weep for the fuel that it adds to the fire of ignorance.
What truly appears to be the case to me is this. To construct a cladogram, the taxonomist needs to be very familiar with the plants and have a good understanding of the characters being used in the process; adjudging them according to some scale of advanced or primitive. The DNA sequencing method requires no knowledge of the plants at all and a phyllogram simply follows a statistical analysis of equivalence of the base pairs. What I see is DNA derived products that so far tell me no more than I already know about the genera, and nothing at all about the species other than some serious mismatches.
I argue for a species definition, not as some vague statement that species are things into which a genus can be divided, or simply a list of names under a Latin genus heading, or the zoological concept of closed breeding systems. My definition drawn from a lifetime of interest, observation and experience in things material as well as esoteric, is that the creation is a conscious one. This is not a new idea of mine. The conflict between religion and science is that the one maintains that creationism followed some sort of seven day event as in the book of Genesis. Whereas science (as we practice it today), has its origins in the quest for freedom from any such religious belief system. Whether creation began as a big bang, and complete or incomplete does not matter to me. We are faced with natural phenomena in the present that we can observe have changed and are changing. I do not think either the cladogram or the phylogram is anywhere near good enough as an answer, because they are two-dimensional diagrams that do not adequately portray the realities of time and space. Not only that, a three dimensional improved tree model will not show how fast things change within a time span. My model of the species tree is a fractal model that suggests species in a process of “chaotic” order. By the word fractal I mean the endless variation that seems to have no order. While I insist that the concept of a species of Haworthia must mean the same as the concept of a species of insect or frog, or bird or mammal, I do not mean that each species of Haworthia has the same branch clarity on any tree depicting relationships.
What I do not know is how fast populations might change in the process of genetic drift i.e. move off in a direction determined just by the local genetic content. Neither do I know how much nor how frequently interchanging of genetic material between populations occurs and how this is affected by distance between them. In the field there is a very distinct flow of difference (or similarity) with geographic spread. I have repeatedly found that with widely separated known populations, new populations between them are predictably intermediate. It does not seem entirely rational that there are not more random isolated individuals. My experience with hybrids is that they are very local to the mother plants while any populations tend to be very confined and local. What is odd is that hybrids may be found despite great differences in flowering times of the parent species.
The reality in Haworthia, however we define species, is that there are populations that are neither one taxon nor another. If the classification were to present a basic requirement that ALL populations known and unknown must fit the model, then definitely we have to consider that there are considerably less species than other writers are prepared to consider. One can continue building a classification from the top in which each new population is simply described as something new and given a Latin binomial. This process was suited to the origins of botanical exploration as well as nomenclature. It does make it a lot easier for collectors and the exciting process of always novelties for the collector. Alternatively one can decide that classification is by this stage a fully predictive one and one that must primarily suit the requirements of botany. If only botanical classification was more professionally and rationally based. Sampling is very extensive and a classification based on the known, is predictive. Personally I am very comfortable that I have achieved this in large measure – not entirely, but close enough and a lot closer than some critics will concede. With all that said and done, it is just a sad fact that formal classification is too often undertaken by people who have a need. That need may satisfy them within the limits of their personal experience and knowledge, with an uncomfortable disregard for the depth of knowledge elsewhere.
Reference: Readers can refer to the article in Haworthia Update Vol.2:149 (2006) “Nectar sugars in the Alooid minor genera and a need for another model”. That article is also posted to the internet with much of my other writing in HaworthiaUpdates.org.
Haworthia mutica (groenewaldii) and its twisted leaves.
In this article Bruce Bayer responds to the notion that apparent leaf twist arrangement is a defining characteristic and further explains his disagreement in recognizing the Buffeljags plants as a new species. (Also see Haworthia flowers – some comments as a character source, Volume 7, Chapter 4:- What is typical Haworthia mutica?, and Volume 7, Chapter 2:- Further exploration in Haworthia. Further to finale.) The species definition Bayer uses is that populations are fractal and DNA is governed by mathematical non-linearity. What does that mean? We have space with two dimensions latitude and longitude, and we have time with two dimensions – calendar time, and the speed (instability) of the arrangement of the DNA base pairs. At any moment in linear calendar time there will be an arrangement of the DNA depending on the stability of the DNA. At one time there will be a clear set of ‘species’ and at another time a different set as the mix of characteristics continually change within and between the populations. ~ Lawrence Loucka
Haworthia mutica (groenewaldii) and its twisted leaves.
By M.B. Bayer
There is some argument about the status of this population of plants at Buffeljags. I have explained my opinion of it based on a species definition that I use. I also have reported on three other populations a short way away at Rotterdam. Furthermore I have discussed the flowers at length in a flower report. These are all available free on Haworthiaupdates.org. Gerhard Marx does not agree. The disagreement first has its roots in what constitutes a species and Marx stays with the standard view that characters are what define species. I opt for the view that species are systems related to geographic distribution and to all the elements that drive vegetation and change (evolution + equals change from some unknown initial condition). I think these patterns of change and difference are fractal i.e. detailed pattern repeating itself. Perhaps it would be more correct to say that the organization of pattern is according to a mathematical function which is non-linear. This means that the end product has many different outcomes. But this is complexity just of argument that none of us can deal with.
What comes into the geographic nature of species is also the nature of habitat. What happens is that we have a set of apparent species in Haworthia with a known distribution range. These species are primarily H. retusa and H. mirabilis. There is clear evidence that H. pygmaea and H. mutica emerge from a milieu of populations of those two and that H. floribunda is also involved. Buffeljags is geographically central to this arrangement and the habitat (wrongly described in the description of H. groenewaldii) is very unlike those where the named species generally occur. It is thus no surprise that the plants appear different. The Buffeljags population and its habitat also differ to a small degree from the west side of the river, but both are essentially geologically fairly recent river alluvial deposit.
Marx is insistent that the plants at Buffeljags are so different as to be a discrete species and I disagree. My disagreement is based on my experience of characters in plants. In Haworthia I think these are few and obscure. Thus it is almost impossible to delineate or circumscribe a species by characters and no one has succeeded in producing an identification key that can work. All the differences of opinion and argumentation about names come down to this issue of a species definition and the characters available to recognize them.
The essential points made for H. groenewaldii as a species is the shiny leaf surface and the flowering time. H. mutica does flower four months earlier – agreed. But H. marginata in the same close area also flowers similarly out of synchronicity with H. marginata elsewhere, as H. minima was also observed to do. In elaborating differences for H. groenewaldii, Marx offered the facts that the plants had fish-tail buds and that H. mutica did not. Very soon after he stated that he did not actually know what the case was in H. mutica.









Fig. 1 to 9, MBB7801 Haworthia mutica (groenewaldii), Buffeljags.
In addition he maintained that the leaves in H. groenewaldii were different as follows…”In terms of the angled newer leaves of H. groenewaldii, have a look at the plants again and you’ll see the young leaves are consistently twisted sideways. A spiraling effect. This never shows up in H. mutica”. I find this statement very odd because such a structural difference would come down to a difference on the level of genus or even family. So I looked at the plants I have in my possession and provide illustrations here to demonstrate no significant deviation in respect of twisting. I have even included a picture of a mature plant (fig. 10) of H. mirabilis to show the same “character”. The spiralling effect is universal in the aloids and is even visible in those species with distichous leaves. In the retusoids, where H. mutica belongs, the leaves have been said to be 5-farious. More usually it is possible to see them as trifarious. In young seedlings the leaves are bifarious as the very basic spiral effect comes into play.

10. 7912 H. mirabilis, Rietkuil
I do not think this is a character one can use to distinguish a species at all. There are many cases where it is fairly possible to characterize populations by a wide range of so-called characters. My opinion is that generally in the many species (by my definition) of Haworthia this can virtually only be done at individual plant level. The Buffeljags and Rotterdam populations are simply the western counterparts of populations around Albertinia (eg H. mirabilis ‘splendens’, H. pygmaea ‘fusca’, H. pygmaea ‘esterhuizenii’) that emanate from the relationship of the prime species that I named above.

11. 7916 H. mirabilis, Van Reenens Crest. 
12. 7925 H. mirabilis, Van Reenens Crest 
13. 7945 H. muticaX mirabilis, Klipbankskloof. 
14. 7967 H. mutica, Spitzkop. 
15. 8005 H. retusa (turgida), Wegwysersrivier.
Comprehension and significance
My writing has been described in all kinds of terms, hermetic and pretentious being two of the adjectives used. Recently Kris Tamayo also suggested that he had trouble understanding what I wrote or write. The fact is that writing and expressing yourself is difficult. But the first place is to be clear about what you want to say. Or is that in fact the first place?
Writing is a means of communication and it really only should start when you are clear about what you think and what you want to say. Then it requires that the listener is clear about what he/she wants to hear and has the common cultural heritage that permits communication and understanding to occur?
In writing and talking about plants I personally get very frustrated by the technical problems of definition and knowledge that mess up communication completely. This is one of the obstacles in classification where there is no species definition and we do not actually know what species are. There are a lot of other obstacles. Recently someone wrote and implied that there were a lot of significant differences among Haworthia that could be used to arrive at a better classification (than any already available). The point I would make is that this person has his own idiosyncratic view of what significance means. This is not strange at all because a prominent scientist was once applying the statistical measure of standard deviation to two and three measured samples. That measure probably cannot be used until many more measurements are made. What is taken to be significant may be quite irrelevant to the actual question of whether there are more or less species. This is why amateurs and collectors should keep clear of classification. The professionals already have too many problems.
The characters we use to make identifications are important in that they may be of the yes and no kind i.e. present or absent, or they may be graded from vague to prominent. So it is very easy to go to one end of the scale and take only the prominent or what happens to strike your eye. This is exactly what happens. Unless followers and interested parties realize the impact this sort of decision making affects what they may want to know and understand, there can never be any harmony and peace in the classification process.
Look at these flowers and see what you can glean from them…



These just happen to be the only three flowers I have of a few plants of Haworthia herbacea from recent sampling. They are shown here in correct proportion to one another with the third being 18mm across at widest spread of the two upper outer petals. So we have two things we can call characters i.e. size and colour that we could say in respect of this simple sample, that they are significantly different. No matter how many times or how we measured these two things, this fact would stay the same. The plants happen to be from two populations and we can then ask if this is true for those populations. I did ask such a question of both H. herbacea and H. reticulata, and ended up by learning that I needed a sample of about 200 flowers to arrive at a statistically true answer at a probability of 95%. The thing is that I could go a little further south and sample another population and get a really pink flower with a spread of 25mm or more that would nearly double the spread of my measurements.
There are several incidentals here. One is the delineation of the mouth into the tube of the flower. Why is it so clear in the third picture? The second is that the first flower has not opened as flat across the face as the other two despite being at the same expected state in respect of time from opening. The third is that the name “subregularis” was used in this genre of flowers because the petals are so equally spread; perhaps less-so in the middle picture. Still a fourth curiosity is that in the southwestern species with the more extreme biarcuate bud with the fish-tail tip, is how the tips of the upper outer petals are “replicate” – i.e. the margins tend to fold together. In the Worcester/Robertson Karoo particularly H. herbacea and H. reticulata have the “regular” flower shape. But in H. mirabilis in this area, the upper outer petals may be held in a plane directly behind the inner outer petal and do not spread at all. There the bud tip is still fish-tail and the upper outer petal tips very replicate.
Habitats

Habitat at Buitenstekloof 
7995 Habitat looking northwards 
KG36-70 habitat viewed from the west. 
View up stream from 2420 habitat 

6631 Habitat.1 
6631 Habitat.2 

Habitat H. floribunda. Plaatjieskop. 
Habitat H. retisa. Plaatjieskop JDV 0711 
Fig. 8a. Habitat of H. elizeae. Stormsvlei. 
Fig. 8. Habitat of H. elizeae. Stormsvlei. 
Fig. 2. A photograph of the habitat of MBB7899. 
13.21 View overlooking habitat 7496. 
Fig. 131 – 7953 N Klipbankskloof westward view of habitat 
Looking to the previous habitat of larger quartz rock 
Fig. 8a. Habitat of H. elizeae. Stormsvlei. 
Fig. 8b. Habitat of H. elizeae. Stormsvlei. 
View further into Buitenstekloof 
Ouhoekberg view to south. 
Ouhoekberg view to southeast. 
Ouhoekberg view to east. 
Ouhoekberg view to north. 
view Hammnsberg to the southeast 
view from Hammansberg northwards 
view from Hammansberg eastwards 
Van Reenens Crest 
View 4 looking northeast from within hiking trail 
View 3 looking northeast from within hiking trail 
View 2 looking northeast from within hiking trail 
View 1 over Greyton from the northeast 

8045 view looking west of north 
View from Brakkloof looking east. 
8048 view looking north 
View looking east of north 
7821 habitat viewed from the west. 
View westwards to the Kanetvlei 
Fig. 49 View from locality MBB7917 to that of MBB 7919. 
Fig. 2 View from locality MBB7914 to Heuningklip. 
Fig. 1 View of locality MBB7914. 
Fig. 23.10 7075 View across site. Grootvlakte 
7075 View of site at Grootvlakte 
Fig. 23 7938 SE Stoffelsrivier View looking east to Sandhoogte 
View looking east 
Fig. 14.14 7497 View looking north. 
13.22 View looking west to north of Kleinberg. 
Fig. 6.3 7928 View looking southwest with Kleinberg off to the right. 
Fig. 6.2 7928 View looking east to 7927 
Fig. 80 7945 View lookin across depression to west facing slope 
7945 View lookin across depression to west facing slope 
Fig. 54 7943 View west to farmhouse 
Fig. 45 7941 View east to Die Kop 
Fig. 44 7941 View west to Haarwegskloof 
Fig. 145 View looking northwest to Riviersonderend 
Fig. 130 – 7953 Klipbankskloof West 3 View westwards 
Fig.100 View looking south to Potberg. 
Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS. 
Fig. 35 View looking south to Tradouw Pass 
Fig. 12 View looking east to Set 1. 
Van Reenens Crest 
Viewing site from the north 
MBB7266 SE Lemoenpoort, view to southwest 
13. A view from south Dwarsindieweg. 
4c. View northwards from Bakoven 
7. View south to Villiersdorp 
1c. View looking west from north Muiskraal 
1b. View looking south west over Muiskraal to Garcia Pass 
1a.View from above Onverwacht North Westward to Ladismith 
View taken looking eastwards from Kleindoorn (Kleindoornrivier) 
View from the upper slopes of the Zuurberg at Wilgerfontein, with Darlington Dam just visible in the distance. 03 . View west of south from Sandberg. View 2 From Cilmor eastwards. View 1 From Cilmor to the northeast. 
Fig. 4 View to the northwest across the flood plain. 
Fig. 3 View northwest from a central position. 
Fig. 1 An overview of the Kruis River Valley. Fig. 14 The typical ‘pressure burst’ of white kaolinic soil from under more solid ferricrete where H. retusa and H. mirabilis are commonly found. 
Fig. 91 The ‘white clay pressure burst’ 
Spitzkop at Riversdale. These flat-topped small peaks and larger tablelands are ferricrete caps often associated with clayish soils on the slopes. They are often the home of Aloe ferox and A. arborescens. Sometimes there are grassy patches and pressure bursts of bare clay. H. magnifica and H. serrata occur in this kind of situation. However, these ferricrete sills are also north of the Langeberg mountains, where species like H. emelyae and H. bayeri may occur. 
8053 looking at Rooivlei 
KG36-70 looking northwards 
7821 looking northwards 
Looking from Swartrug towards Riversdale. 
Looking from Morning Star to Kransrivermond 097 
This is a picture taken from a very late flowering population of H. maculata near Die Nekkies resort south of Worcester. Looking north to where H. maculata grows at about 4000ft on the Audenberg peak, and to the right at about 1000ft in the lower Hex River pass. That is where it occurs close to H. aff nortieri. 
Fig. 90 The countryside looking towards the Tradouw Pass. 
Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – b 
Fig. 3.7 Looking west to the Kleinberg 
Also looking SE towards Die Kop and Potberg 
Looking SE towards Die Kop and Potberg 
MBB7266 SE Lemoenpoort, looking up at locality. 
5. Looking north with M marking the location of Haworthia maculata, K – Karoo Garden 
4. Still further east looking west northwest 
2. Same spot looking east northeast 
1. Looking down the north face Die Nekkies east to west 
1e. Looking southwest to Garcia Pass from same spot as above 
1d. Looking eastwards over Muiskraal
MBB6694 Kanetvlei, Hex River Valley as a variant of Haworthia nortieri.
In Haworthia Update Vol. 9 dealing with H. maculata, I again draw attention to several populations that are problematic in an area very difficult to explore. The populations are MBB7865 at Keurkloof southeast of DeDoorns, EA1441 at the Hex River Pass, an unrecorded population south of Sandhills (about 3km east of Kanetvlei) reported by Ernst van Jaarsveld. In addition there are two populations apparently of H. arachnoidea at Tunnel Station east of Osplaas.
The illustrations that accompany this article are:
- MBB6694 Vreesniet, Kanetvlei.
- EA1441 Hex River Pass, east DeDoorns.
- MBB7865 ex E. Van Jaarsveld, Keurkloof, DeDoorns.
This population MBB6694 is at Kanetvlei southeast of Sandhills, and I originally ascribed it to H. arachnoidea but this is not correct. To this I can add unexamined populations reported by PV Bruynsdeep in the Langeberg at Keerom Damand another of Ted Oliver in the mountains north of Nuy. MBB6694 is only 200-300m north of MBB7994 also in sandstone. The sandstone strata are by no means all the same and one can even find shale bands in what is essentially a sandstone formation. H. maculata flowers late winter to early spring and these other populations in early summer.
View westwards to the Kanetvlei
I have not examined the flowers in any number and remain doubtful that this information will clarify any deductions that can be drawn from the vegetative characters or the geographic positions of the populations. It appears to me that consideration must be given to a relationship with H. nortieri that in my assessment occurs from the far northwest to as far east as Prince Albert {i.e. H. nortieri (devriesii)}. What I have seen of the flower, and shown now is that it definitely does not belong in the context of the Southern Cape species where the bud-tips tend to be “fish-tailed”. The inflorescence are slender and few flowered.
There is some criticism of my emphasis on geographical distribution as a key indicator of species. It was even said, as though I would have denied the fact, that I could not identify plants unless I knew where they came from. There is an incredible amount of both ignorance and the obtuse behind these statements. One critic feels that I have ignored detail and even ignored flower characters in the development of my opinions. This same critic claims a high degree of agreement with me (80%) based apparently on the view that so much of my classification is correct (meets with his approval), and presumably that the 20 percent that he contributes to the system makes it 100% correct. This is wholly wrong. This critic is simply unable to bridge the historical chasm between his introduction to, or knowledge of, Haworthia and mine. He simply does not recognise the shift in method and why I switched from a “character-based” approach to a phytogeographic one. It is really very simple in that the character based system as used from the very beginning of classification, has not produced a solution to the identification and classification of Haworthia. It all rests on a very weak and shaky definition of species determined by breeding barriers and consequently that morphological differences necessarily exist. My approach was to show that breeding barriers (at least in plants) could be integral to species and that morphological differences could equally be nothing but variation within species. Therefore I arrived at a definition of species from the view that they were phenomena spread in space and changing with time. This is just a fundamental of plant species as they constitute the vegetation and the floras of the world. Virtually the first question following “What is that?”, is “where does it come from?”.
Why 6694 is so interesting is precisely because it occurs in an area which is so poorly known and represented in our knowledge of Haworthia. The reality in the subgenus Haworthia is that there are several areas of great species richness and there is real pattern in the distribution of the various elements as they are listed in the 80% agreement area. The 20% disagreement zone concerns truly trivial opinion and unsubstantiated statements, and almost ignores the real reasons for disagreement at all. The real reasons are the realities that superficial morphological difference does not mean species and I have posted a vast amount of material that demonstrates that. I have shown repeatedly that differences in single characters in what can be rationally said to be the same species, can be greater than between different species.
In the case of 6694 I did not pick any particular character to identify the plants as H. arachnoidea from probably as far back as 1975 and I can find no earlier record of this location. I do recall a specimen in the Compton herbarium made by W.F.Barker that I mentally allied with H. arachnoidea but it is not cited as such in my revision. The identification was based on little else other than the spiny-ness of the plants and the fact that the nearest known locality for that species was a very new one of mine a few kilometres to the southwest. At the time, H. nortieri was barely known and this was from far to the northwest at Clanwilliam. Since then H. nortieri has become much better known although still some distance north of Ceres, and with its globose-flowered variants at Laingsburg and eastwards. H. arachnoidea remains remarkably unknown from the Cedarberg and Koue Bokkeveld mountains but does appear in the upper Hex River Valley. Here it is odd that the leaves tend to have the translucent dots that one would expect to characterise H. nortieri. But otherwise the leaf coloration is the much darker green that is associated with H. arachnoidea. A twist is that at Kunje, southeast of Citrusdal, H. nortieri does have very dark green leaves and there is no doubt that confusion with H. arachnoidea is inevitable. I have very little to offer in this regard because this degree and level of confusion or doubt is intrinsic to the genus anyway.
Why I now decide to relate this Kanetvlei population to H. nortieri is not to be construed as a decision now to call white what previously was clearly and definitely black. It is taking all my knowledge and experience to suggest simply that this is a better reflection of the situation where there is not enough hard data to determine identity at a higher level of certainty than about 50%. What I do suggest that there are many situations in Haworthia as the case with H. maculata and H. pubescens only recently exposed. In this case the two very different elements can be very confidently be said to be the same in respect of distribution and linking populations AND characters. Throughout, particularly the subgenus Haworthia, we have situations of continuity and spatial (geographic trend) and nearly all my many publications deal with exactly this reality. What it suggests is that my critic should come to consider if he has some other motive between punting points of difference that prevent him from finding points of agreement, besides those that rest coldly on logic.
1. MBB6694 Vreesniet, Kanetvlei.





























Flower profiles.









Flower faces.









Flower buds.



2. EA1441 Hex River Pass, east DeDoorns.


















3. MBB7865 ex E. Van Jaarsveld, Keurkloof, DeDoorns.








Haworthia minima and pumila flowers

This flower (H. pumila) is apparently persistently regarded by botanists as actinomorphic (star-shaped, radially symmetrical) – as though zygomorphy (yoke shaped, bilateral, asymmetrical) in the aloids is an uncommon condition!
Radial symmetry means the flower can be divided into 3 or more identical sectors which are related to each other by rotation about the centre of the flower. Typically, each sector might contain one tepal or one petal and one sepal and so on. It may or may not be possible to divide the flower into symmetrical halves by the same number of longitudinal planes passing through the axis. Zygomorthic flowers can be divided by only a single plane into two mirror-image halves, much like a yoke or a person’s face.
If you see the way the inner upper petal overlaps BOTH the two lower inner petals, you recognise that there can not be actinomorphy in aloid flowers.
Haworthia pumila








Haworthia minima















MBB7989 Haworthia pumila, Lemoenpoort
Kobus drew my attention to a glabrous plant of H. pumila while he was photographing this species at Lemoenpoort. The plants here have a missing chromosome and tend to have a purplish colour. I have seen smooth non-tubercled leaves elsewhere. But do check out that one plant – if you look carefully you can see the leaves are in 8 nearly vertical tiers. Proper botanists recently, for Taxon, described the arrangement of leaves like this in only two tiers (e.g. H. truncata) as “distichous insertion”. This is weird. Are the leaves in this plant of H. pumila “octichous”? Is H. viscosa “tristichous”. No. The leaves in the aloids are alternately and spirally inserted.


















Flower profiles



















Flower faces
















Leaf arrangement in Aloe striatula
Herewith is an image of the leaf arrangement in Aloe striatula. I have numbered the leaves in inverse order to show that the leaves are as much distichious as trifarious i.e in two rows or three. The primary set is 1 through 10, the distichous set is 1, 3, 5, 7, 9 and 2, 4, 6, 8, 10. The trifarious set is 1, 4, 7, 10; 2, 5, 8 and 3, 6, 9. I have also added a picture of the leaf insertion – here it is entire and there is a leaf sheath around the stem – the actual point of insertion is at the pointer although the “veins” continue through that point to the stem itself. In this species the next leaf is inserted just below the opening of the previous sheath. Despite the leaves being alternate, they are spirally arranged. In Aloe broomii the leaf insertion is continuous and you can peel all the leaves off the stem in one piece. In most Haworthias the leaves are imbricate (overlapping alternately) but always in a spiral sequence. H. wittebergensis has leaves that have entire insertion and by memory the same is true for H. blackburniae and maybe for H. viscosa.


Just what do we do with names for Haworthia?
Previously published in BCSJl (Cactus World) 30.4:211(2012)
M B Bayer
Taxonomy always provokes differing views, and Haworthia in particular has been subject to years of vacillation. The author has long been a campaigner against the haphazard proliferation of new names for every new, morphologically different population or variant. He questions the vagueness of a conventional species concept and pleads for a more reason-based, logical and sensible, communal approach to understanding and classifying Haworthia species. He hopes in this article to convey a message which is relevant to whatever genera of plants you grow. Photography by the author.
In Haworthia, professional botanists have struggled with and avoided the group because it is so infused with amateurs whose interest in the genus far outweighs any knowledge of botany. So classification of Haworthia has muddled on with scant regard for the discipline of botany as a science. Whether that has changed, is not for me to say. I have personally been gathering information on distribution and variation for over 50 years and have very seriously tried to keep that in the context of the science I was trained in and present it in a manner that botanists can follow and hopefully accept.
What has developed is that academic and professional botanists have been working with the tools of molecular biology and the results of five independent studies have all pointed towards the same conclusion, which is that the species of Haworthia are elusive, and the related genera are also not adequately distinguishable in the DNA data. The best solution they can offer is to merge the genera back into a single genus, namely Aloe. My personal reaction is that this is not a new idea and also Gordon Rowley pointed this out as far back as the 1970’s. I have also said that the subgenus Haworthia does not sit comfortably with the other two subgenera. Had I been a true taxonomist I would have implemented that by separating Haworthia into three separate genera and that is what I really would like to see. But the reality is that this does not solve the other problems that exist regarding Astroloba, Chortolirion, Poellnitzia, Chamaealoe, Leptaloe, Lomatophyllum, and the small Madagascan aloes. True botany alone can resolve the current and a new classification (still in manuscript form) has been proposed by a group of scientists that will really ‘rock the boat’ as far as collectors are concerned.
In Haworthiad (2012:4), I wrote about Haworthia mirabilis ‘submagnifica’. (Ed. note: The name in inverted commas, according to the author’s system indicates a variant name rather than a formal conventional variety, subspecies or form. It is his contention that there is no species definition and thus formal names have a large element of uncertainty. Another option is to drop the use of any rank denotion at all). It is one of the first populations linked to von Poellnitz’ H. magnifica long before so much was learned about distribution and variation. I used the prefix “sub” because this means “somewhat”, “almost”, “slightly”, “partially”, and possibly a few other words that mean… it is, but it is not. The population concerned is Komserante (Figs 1-2) and this particular population has acquired the name H. vernalis (Figs. 3-4). But I think we need to start from scratch and drop all the ‘baggage’ of the years. I personally have learned so much since I wrote my revision in 1996 that I know it is not possible to properly backtrack and retrace the passage forward by the use of Latin names. Interfertility is the basis of the system we use to identify and describe species and my field experience was already proving that this cannot possibly apply in the way in which Haworthia has been, or is to be, classified.


It is quite evident that H. retusa and H. mirabilis, both of which I accept in a very much broader context, do hybridise and there are populations that fit between. But first let me just explain that I regard H. retusa now to include H. turgida and all the variants of that species (nomenclatural priority obviated the use of the name turgida to cover the greater body of populations for this species). In the same way I regard H. maraisii, H. magnifica, and H. heidelbergensis and whatever variants were attached to those, as H. mirabilis. There are thus two species. My further observation is that H. pygmaea and H. mutica are segregates from the common gene pool of H. retusa and H. mirabilis. The Komserante population is the one in which that same gene pool is re-combining (Figs. 5-10). I am not in the least sure of all the intricacies but it seems to me that it is actually the group of populations that I recognize as H. retusa var. nigra that is pivotal in the relationship of all these species that I recognize. In this we discuss populations using names as prescribed by convention. This convention caters for chronology and authorship and not for evolutionary pathways. Both the names “retusa” and “turgida” precede the name “nigra”, but the populations that I now assign to H. retusa ‘nigra’ may better fit the concept of evolutionary origin.






The picture is complicated by the role of H. floribunda. This also hybridizes with both H. mirabilis and H. retusa as odd hybrids as well as at a population level. Just what are we to do? History has demonstrated all too well that a bevy of ill-assorted interested parties trying to impose a botanical classification is going to produce nothing but conflict and confusion. This has been going on now since the time that Smith, Von Poellnitz and Resende were simultaneously describing new species. How are we going to turn around and arrive at the understanding and stability of names that we seek?
In the past I have been extremely reluctant to make the following suggestion, and even now am a bit hesitant. What we need to do is turn to people who are employed to do this work. Herbaria and herbarium botanists are tasked and entrusted to classify and name plants. Perhaps it testifies to the complexity of the subject, or just its enormity, that these botanists have too often needed to defer to amateurs who have the interest, energy and enthusiasm and commitment to acquire field knowledge that a professional could never get the time or funds to do. The unfortunate part is that sometimes amateurs may not be able to relate their knowledge adequately to academic botany.
So what is the solution? It is that the community leaders assume the responsibility for the establishment of a system of classification that is meaningful to the community they serve. (By community, I mean initially the botanists who should be providing us with scientifically sound classifications, then editors who are familiar with what botanists do, next are the Societies with their memberships, and also opinion formers in those groups. Finally included are the reviewers and commentators who lead opinion in one direction or another). It is surely not that difficult to sit down together to discuss and arrive at a set of guidelines by which a decision can be reached as to what (not whose) system to accept. A solution does not belong to anyone. Latin names are assumed to refer to an entity called a “species”. Botany has indeed been very lax and remiss in not providing a definition and this is the prime reason why amateurs have had so much freedom in generating Latin names for the most frivolous reasons. Botanists themselves have often not been far behind.
I think it is time to change all this and must excuse myself from any decision making body or process because I have a vested interest in respect of all the words I have written on the subject.
M B Bayer
PO Box 960
Kuilsriver 7579
South Africa
Email: bbayer227@gmail.com
Literature cited:
Bayer, M B(1999) Haworthia revisited. Umdaus Press, Hatfield.
-(2012) Plants in my collection 7: H. mirabilis ‘magnifica’. Haworthiad 26(1): 4-5
The absurdity of taxonomy and nomenclature?
In Alsterworthia 13.1:6 (2013) there is yet another statement about the correct name for a species of Haworthia. It reads …”The vexing matter of the correct name for Haworthia pumila has taxed some of the finest minds in botanical nomenclature”. The article then goes on to replace that name with H. margaritifera with an explanation so simple that it casts considerable doubt onto the quality of those minds that have examined the problem. There is of course also a difficulty in that the quoted sentence implies that H. pumila is the correct name, while the article goes on to dismiss it.
The fact is that Linnaeus listed four different things (varieties) under a single name Aloe pumila, and the only issue was about which of those four things ends up with that particular first name. It so happened that Burman in 1701 made a choice, and was followed by Aiton in 1789 who chose something else. So the name Aloe pumila stood but applied to two different things (species). Duval was unaware of the earlier Burman choice and used Aiton’s choice when he created the genus Haworthia. Dr L. A Codd, whom I would have accepted as a fine mind (and also as a very ethical man), advised C.L. Scott that Aiton’s choice was in fact illegitimate and hence also Duval’s usage in Haworthia. The opinion was that Burman’s choice was the first and also thus the legitimate one; and it could not be denied by the illegitimacy of the Aiton usage as a later action. So perhaps it was/is a question of rank or just an opinion that Aiton made up pumila as an entirely new name.
There is nothing complicated and mind boggling about this simple state of affairs. Or is that so? What on earth does the ICN as the product of presumably fine minds actually say about this? Does it take 50 years of debate to establish such a simple fact? The situation is further exacerbated in that the finest taxonomic minds are involved in an epic battle to either create a single alooid genus or many lesser genera. It appears that the latter option is winning ground although the war against single-species genera has surely not been abandoned. When the dust finally settles, it will be recognised that the taxon onto which this unfortunate species viz. Haworthia pumila/margaritifera resolves, will be a separate genus, probably Tulista, and then the truly correct name will be Tulista pumila. Or will it?
My personal opinion that however the case may truly be judged, the correct answer is the intent. Scott and Codd came to a workable end point way back in history and it has been my misfortune now to have defended that. I think there is a parallel in the case of Aloe bainesii. Put into use by Reynolds far back in history, it is found that the name barberae had page priority and thus preference. In what interest was the change made? Why does the code have a conservation facility for names? The fun seems to be in the argument rather than in usage.
The argument that I think L.A. Codd would have made is this. There are four varieties covered by the Linnaean epithet pumila. The first effective use of that epithet for one of the four was to the warty t10 of Commelin and that is how the name is formally typified. To use of that same epithet at any other time for any other of those four Linnaean varieties would be illegitimate. It is also not in the least certain that the name margaritifera is correctly typified by Wijnands on the same Commelin illustration t10. I aided Wijnands to this conclusion before myself stumbling on the fact that its correct typification would be on a Bradley illustration.
There is a curious twist to the issue and it is somewhat of an oversight that the persons involved never read the introduction to Haworthia Revisited. I explained the problem and also in respect to the correct application of the original name margaritifera to what we know as H. minima. I also cited the name H. pumila with the authors as (L.) Bayer to make it clear that I was not accepting the name with the authors (L.)Duval as Scott cited it. I thought at the time that it was a mistake on Scott’s part. I had also written to Dr Codd specifically about the issue and this is when he explained to me (as a professional taxonomist and fine mind) that the illegitimate use of the name pumila in Haworthia did not prevent the correct usage. It only strikes me now that he probably had advised Scott to the effect that the NAME as Duval had taken it through to Haworthia was correct, even if he had applied it to the wrong species. This whole issue has not been properly and fully aired. To argue that it is a new name seems to me just a piece of intellectual vanity that serves no purpose other than to demonstrate our collective failure to honour the intent of the code – or respect the dismay of interested person. A last point I make is that people can and will always find topics to disagree on, so this is an important trap to avoid and be mindful of. It is not particularly in the case of nomenclature that this seems to happen. I had no doubt at the time when I made the decision to accept Scott’s usage that no matter what my decision was, cause would be found to change it. If I had decided on either margaritifera (correctly typified) or maxima (as I. Breuer later did), this would also have been argued as wrong.
[-ed. There seems to be a number of taxonomic changes brewing. Time will tell whether pumila survives.]
References:
1. Haworthia margaritifera/pumila
Dr. John Manning. S.ANBI.
The vexing matter of the correct name for Haworthia pumila has taxed some of the finest minds in botanical nomenclature. Since I do not include myself among their company, I was not in the least surprised to find that I had misrepresented the situation. Thanks to expert input from Roy Mottram and Urs Eggli we can now put the matter to rights.
The issue of the correct name for Haworthia pumila starts with the fact that in his original publication of Aloe pumila, which forms the basis for this species, Linnaeus recognized several varieties, but without explicitly listing the typical variety, thus he did not list Aloe pumila L. var. pumila. Linnaeus’ Aloe pumila was subsequently effectively lectotypified by Burman f. (1701) [and later in enor by Scott (1978)] against the illustration in Commelin’s Horti medici Amstelodamensis, which is also the type of var. margarit!fera. This renders the name margaritifera homotypic with Aloe pumila L. (i.e. they share the same type). As the autonym (i.e. following automatically from the species name) for this species, pumila would normally have statutory priority over margarit(fera BUT, in the interim, the combination Haworthia pumila (Aiton) Haw. (1804) had been published, based on the name Aloe arachnoidea var. pumila Aiton, a quite different species that we know now as H herbacea. The publication of this combination renders Haworthia pumila (L.) Duval (1809) an illegitimate later homonym and thus not available for use in Haworthia. Because the combination Haworthia pumila cannot be used for Aloe pumila L. as a result of its prior usage for some other taxon it must be substituted with the next available valid and legitimate epithet, which is margaritifera. Note, however, that in any genus other than Haworthia the epithet pumila is the correct one to be used for this species.
The formal rendering of this situation is as follows:
Haworthia margaritifera (L.) Haw. (1819). Aloe pumila var. margaritifera L. (1753). Aloe margaritifera (L.) Burm.f (1768). Aloe pumila L. (1753). H pumila (L.) Duval. (1809), hom. illegit. non H pumila (Ait.) Haw. (1804). Lectotype, effectively designated by Burman f. in Prodromus florae Capensis: 10 (1768) [Superfluous lecotypification by Scott (1985)]: Illustration in Commelin, Horti medici Amstelodamensis, Pars altent: t.l 0 (1701): Aloe Afric: folio in summitate triangulari margaritifera, flore subviridi.
2. International Code of Nomenclature for algae, fungi, and plants (Melbourne Code)
3. Aloe pumila, Haworthia pumila; what or who is confused? ISBN: 0-9534004-4-1 Bruce Bayer. Alsterworthia International Special Issue No.3. https://haworthia-updates.haworthia.org/aloe-pumila-haworthia-pumila-what-or-who-is-confused/
4. Commelin, Johannes, Horti medici amstelodamensis rariorum tam Orientalis, vol. 2: t. 10 (1701)
http://plantillustrations.org/illustration.php?id_illustration=122192
5. Commelin, Johannes, Horti medici amstelodamensis rariorum tam Orientalis, vol. 2: t. 11 (1701)
http://plantillustrations.org/illustration.php?id_illustration=122193&height=750
6. Curtis’s Botanical Magazine, vol. 33: t. 1360 (1811) [S.T. Edwards]
http://plantillustrations.org/illustration.php?id_illustration=8348
7. Moninckx, J., Moninckx atlas, vol. 3: t. 12 (1682-1709)
http://dpc.uba.uva.nl/cgi/i/image/image-idx?c=botanie;view=entry;cc=botanie;entryid=x-421058064 and
http://plantillustrations.org/illustration.php?id_illustration=133655
8. History of Succulent Plants, Bradley, Richard (t30) (1716)
http://www.botanicus.org/page/614136
10. History of Succulent Plants, Bradley, Richard (t21) (1716)
http://www.botanicus.org/page/614116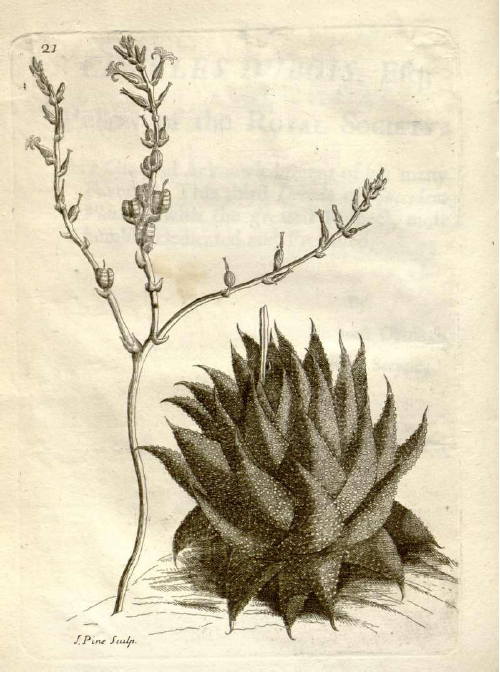
Ed. – another …
11. J., Moninckx , Aloe Africana, folio in summitate triangulari / Margaritifera, Flore subviridi. / C: Commelin, Hort: Amst: Part: 2. pag: 19. Wijnands, D.O., The botany of the Commelins, Rotterdam 1983, p.134
http://dpc.uba.uva.nl/cgi/i/image/image-idx?c=botanie;view=entry;cc=botanie;entryid=x-421058064

Variable chloracantha
Variability – sounds so hackneyed now but still it does not seem as if anybody “gets it”. These are plants from two populations of chloracantha east of Herbertsdale. Two very different soils. One is shale and I show only a single plant from there – the left one of the three unearthed plants. There the plants were “ordinary” chloracantha. But the other locality was exposed “pressure burst” kaolinite such as you get around Riversdale and westwards. Some of the plants reminded me of floribunda and even parksiana. I know of only 6 populations around Herbertsdale and there the plants are different too.












Leaf sequence in Haworthia emelyae ‘comptoniana’ over a long period

1 December 2011 
16 July 2012 
15 September 2012 
13 December 2012 
13 April 2013 
August-2013
Here is an interesting series of pictures showing Haworthia leaf replacement over only 16 months. The plant is in an outdoor rockery. Leaves are numbered from oldest to youngest. Leaf loss has not been quite sequential as the 7th leaf outlived the 5th. In the growth condition of September 2012 the leaves show three tiers viz. 1,4,7,10,13 / 2,5,8,11 / 3,6,9,12. The figure for Dec 12 illustrates the “5-tiered” artifact of the spiral sequence with leaves 15/10, 14/9, 13/8, 12/7 and 16/11/6 in each tier.
These pictures demonstrate that the plants grow quite fast. They may not be very long-lived in nature. I estimate a life span of 15-20 years?
The first picture of this plant was taken in Dec 2011 when the leaves were numbered 1 to 10. Here in March 2014 leaves 1 through 12 have died away and there are 10 new leaves. At the same time the stem will be stretching as roots are replaced in a similar sequence. The life of the plant is going to be determined by the way in which the old stem and roots decay and how that decay will affect the ultimate health of the whole plant.

In the field it seems as though the decay of the old leaves and stem is well regulated as a sort of dry composting. In cultivation this can vary from this state to when all the roots and even the base of the stem can rot unhealthily. In the field one does finds moribund old plants that are at the end of their life and with large aged stems. Just how long the plant can maintain good health in either the field or in cultivation is an unknown. A grower just has to depend on experience and feel for his/her plants and this is why some time back I guessed at the normal life of one of these ‘retuse’ haworthia at about 10-15 years. Some things like T. pumila and H. coarctata are quite different as the stems can elongate and re-root along the ground. Plant of H. coarctata more than 3meters in diameter must be very old. Even H. reticulata or H. herbacea do this – look at H. cymbifomis ‘ramosa’. What you can consider is that new leaves and flowers follow a regular process and it is useful to remember that the plants go through a non-growing stage in the year. I tend to think most haworthia grow in the spring and autumn disliking the heat of summer and the cold of winter. But this is just a generalization that points to things that need to be considered. All species are not the same. Good root health is also associated with drainage and adequate air space in the growing medium.
In species like coarctata and reinwardtii – and I doubt that these are two different species – the plants easily lose obvious connection to the mother clone and a single clone can come to occupy a very large area. H. cymbiformis and H. angiustifolia can also spread widely by pieces breaking off to establish somewhere else. H. limifolia does it by stoloniferous expansion as can H. marumiana and H. zantnertiana. But coming back to the solitary non-pupping growth forms it would be interesting to know just where the essential life of the plant resides. Sometimes the stem just seems to get too thick and cutting it back too far also cuts away the support and source of the very young and new centre leaves.