Incidentally both the soft shale bands within the Sandstone strata as well as the Bokkeveld Shale above, decompose to kaolinic clay.
I took so many pictures that I will just post because I was so fascinated myself. The population is in an area quite new to me and wholly empty as far as haworthia records go – but perfectly predictable if you throw out the chaff. In the meantime, keep thinking.
The plants were all in much the same situations among sparse restioids and grass and very few other succulents. Next I will post better pictures of a few plants and explain a bit more.
What do we have so far? Mirabilis, ‘magnifica’, atrofusca, pygmaea, comptoniana, esterhuizenii, Quite good actually – but some serious omissions. Any more offers – these are all excellent.
Just to make sure the name “magnifica” makes sense, here are a few pictures taken at the type locality.
While looking at those magnifica pictures I was struck by these two to show how a retusa influence is present.
A closer look at the new find.
The scenery
Those flat top hills are the remnants of an ancient african plateaus prominent from near Barrydale to beyond Uniondale – north of the Langeberg. – silcrete upper layer like the concreted layers also present topping the inselbergs south of the Langeberg.
So what is it? If H. magnifica makes sense to anyone as a species, that person is lost to me. This new population is in the sequence from Muiskraal eastwards and it is H. emelyae (‘breueri’, ‘wimii’). The identifications put forward are nearly all in the context of H. mirabilis which includes ‘magnifica’. This naturally supports my perceptions that H. emelyae is in fact the karoo extension of H. mirabilis. More widely that H. mirabilis, pygmaea, retusa and emelyae form a single system. If one reads in the Updates the reports of Towerlands/Aaasvoelkrans one will appreciate the connection better. The “magnifica’ as a species myth is simply and easily dispelled by studying the floral data presented in updates. There is no reason why there should be any doubt and confusion that the cabal so enjoys and wallows in. Aloe barbara jeppeae indeed.
The truth of the matter is that there is no taxonomic solution other than recognising this and stop prostituting science in the name of populism. There are hundreds if not thousands of people interested in Haworthia, and millions affected by the formal names of plants. I see some responsibility in respecting this fact and even more so in trying to understand what all this marvelous stuff really means. It comes down to what a species truly is and how it is defined. If I could bring just one person to that realization I might feel comfortable – with no one caring a damn, I am not. :) I have not far or long to go now and I will be posting more on this topic before I am finished.
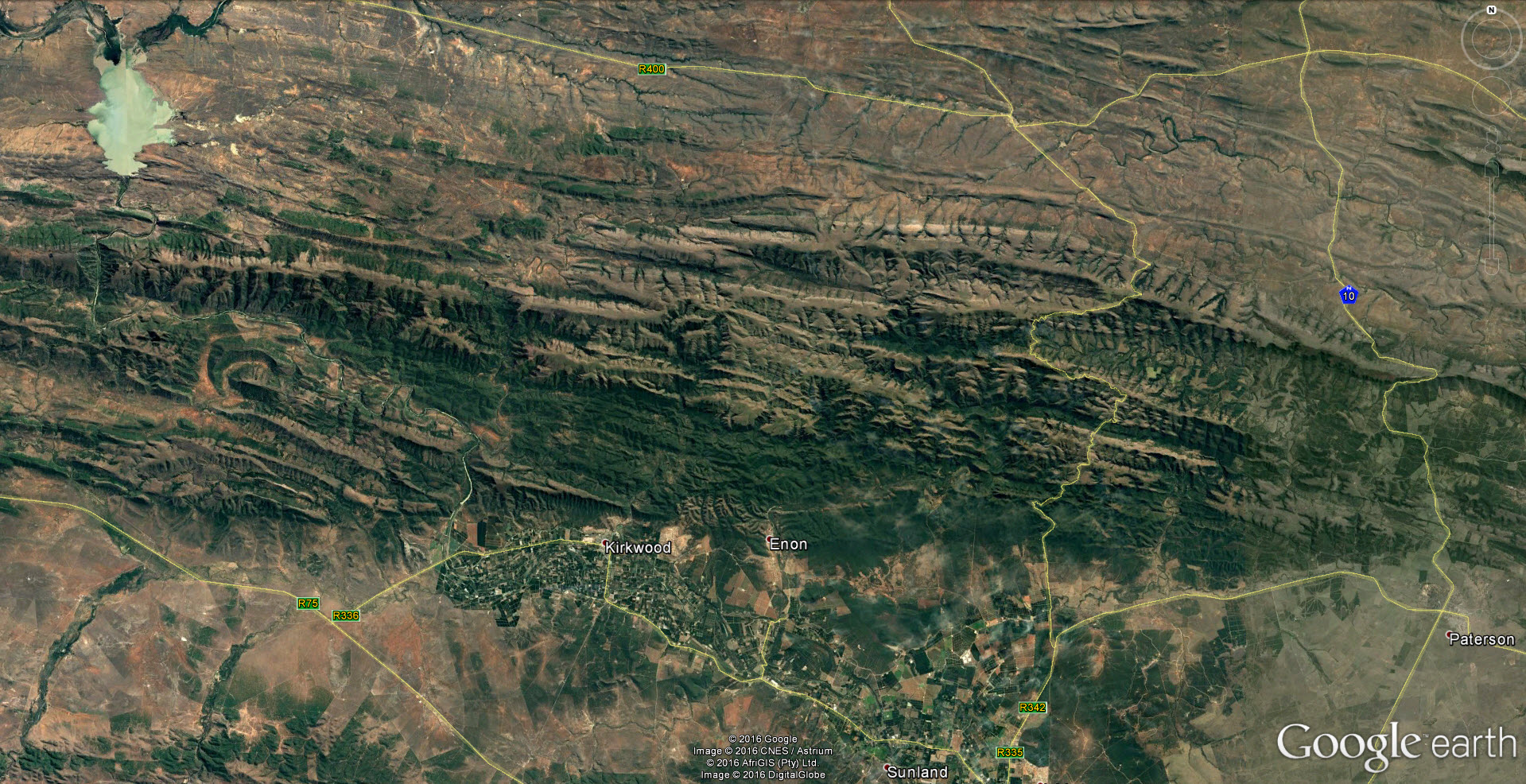
Kaboega is located in the Zuurberg Mountains north of Port Elizabeth in the Eastern Cape province of South Africa. Read more about this in Haworthia Update Volume 1, Chapter 5:- The Haworthias of Kaboega. There are a mind-boggling array of Haworthia populations here in an area considered to be the meeting point of several vegetation biomes. There is much exposed rock, and the soil is very skeletal, composed of three major groups: sandstone, mudstone, and glacial deposits. These pictures are of a Haworthia cooperi variant that occurs high up on sandstone. I went to this spot because researchers had sent me a picture of a cycad festooned with Haworthia. I did not get to the exact spot but have seen the way it forms hanging bundles in other situations.
Haworthia glauca!! can also be found here. On Kaboega these plants often have a very close resemblance to H. coarctata and it is no co-incidence that the distributions of these two species complement each other. An essential element of species recognition is their juxtaposition and if they occur in very close association or not. Darwin said as much.
I visited four populations of this greenish cooperi. One can find plants like this from east of Grahamstown right through to the Little Karoo. Here they are on Dwyka (glacial) skeletal soil.
These next are in the shales low down in the valley on Kaboega – I name it H. aristata. It is very common in the area but complements H. cooperi while there are populations that are neither. Populations cannot be treated in isolation and there is a distinct possibility/probability that I have been too generous with species. The attempts to find answers via DNA sequencing should make the vendors of that technology thoroughly ashamed.
More of these green things. I would guess that these would class as the simple progenitors of cymbiformis and cooperi. Perhaps even of mucronata?
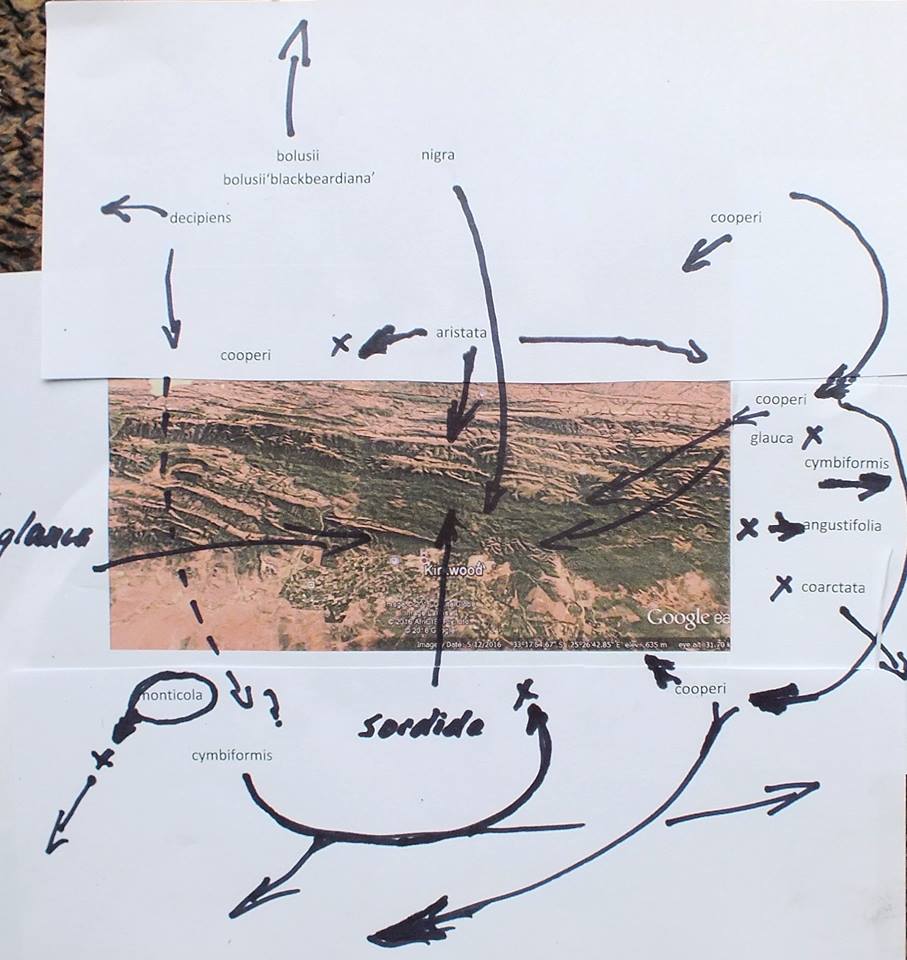
This is Haworthiopsis sordida that does not occur, as far as I know, north of this. H. nigra also occurs here at it’s most southern at this longitude. Altogether it is quite a complex network of distribution patterns that relate to greater plant geography.
From another population as variants on a theme. have seen about 30 such just on this small mountain area and it just suggests what is still unseen on the length and breadth.
Not a great diagram but a way to appreciate the drammatic choreography of plant distribution and how it impacts on classification. Without it Haworthia names make no sense other than as imagined and fantasized. Cooperi and cymbiformis occur as intertwined species to the east and south. In the south they extend westwards to get lost in H. mucronata. Cymbiformis as an independent species does not enter Kaboega except as an observable variant of H. cooperi. The cooperi gets lost westwards as variants of H. decipiens. Perhaps close northwards as H. aristata. H. glauca does cross the Zuurberg but is here confused with H. coarctata that may occur in recognosable form on the eastern tip. Angustifolia is on the eastern end too but does not enter Kaboega. Neither do H. monticola or H. zantneriana from the west. This is also closely tied to the intrigue of winter vs summer rainfall and still further to the massive geological changes of the very recent..
7865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 005
7865 H. cf. arachnoidea. Keurkloof, SE Dedooorns 006
H. cf. arachnoidea. Keurkloof, SE Dedooorns 008
7995 Habitat looking northwards
View 4 looking northeast from within hiking trail
View 3 looking northeast from within hiking trail
View 2 looking northeast from within hiking trail
View 1 over Greyton from the northeast
8053 looking at Rooivlei
8045 view looking west of north
8050 general environment
8048 view looking north
KG36-70 habitat viewed from the west.
KG36-70 looking northwards
View looking east of north
7821 habitat viewed from the west.
7821 looking northwards
View up stream from 2420 habitat
Fig. 3 Below locality at fig 2, southward.
Fig. 2 Westwards towards homestead.
The Skeiding homestead
Looking from Swartrug towards Riversdale.
Looking from Morning Star to Kransrivermond 097
View westwards to the Kanetvlei
Habitat H. floribunda. Plaatjieskop.
Fig. 14 The typical ‘pressure burst’ of white kaolinic soil from under more solid ferricrete where H. retusa and H. mirabilis are commonly found.
This is a picture taken from a very late flowering population of H. maculata near Die Nekkies resort south of Worcester. Looking north to where H. maculata grows at about 4000ft on the Audenberg peak, and to the right at about 1000ft in the lower Hex River pass. That is where it occurs close to H. aff nortieri.
Habitat H. retisa. Plaatjieskop JDV 0711
Pienaarsrivier – truffeling.
EvJ19550.a Blinkberg.
The nortieri plants are just at the small distant upper notch in the sky-line.
17b. Terrain south of Kunje.
Fig. 8a. Habitat of H. elizeae. Stormsvlei.
Fig. 8. Habitat of H. elizeae. Stormsvlei.
Fig. 106 Silcrete hilltops
Fig. 90 The countryside looking towards the Tradouw Pass.
Fig. 2. A photograph of the habitat of MBB7899.
Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – b
Fig. 49 View from locality MBB7917 to that of MBB 7919.
Fig. 2 View from locality MBB7914 to Heuningklip.
Fig. 1 View of locality MBB7914.
Fig. 27.12 7934 Wolwefontein
Fig. 25.16 7937 N Platkop SE Langvlei
Fig. 25.15 7937 N Platkop SE Langvlei
Fig. 23.10 7075 View across site. Grootvlakte
Fig. 23 7938 SE Stoffelsrivier View looking east to Sandhoogte
MBB7935.12 Langvlei The plants are in a tiny bit of rock just beyond the dumps.
Fig. 14.14 7497 View looking north.
13.22 View looking west to north of Kleinberg.
13.21 View overlooking habitat 7496.
Fig. 6.3 7928 View looking southwest with Kleinberg off to the right.
Fig. 6.2 7928 View looking east to 7927
Fig. 3.7 Looking west to the Kleinberg
Fig. 80 7945 View lookin across depression to west facing slope
Fig. 54 7943 View west to farmhouse
Fig. 45 7941 View east to Die Kop
Fig. 44 7941 View west to Haarwegskloof
Fig. 175 JDV92-64 Very localised.
Fig. 145 View looking northwest to Riviersonderend
Fig. 131 – 7953 N Klipbankskloof westward view of habitat
Fig. 130 – 7953 Klipbankskloof West 3 View westwards
Fig.100 View looking south to Potberg.
Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS.
Fig. 35 View looking south to Tradouw Pass
Fig. 12 View looking east to Set 1.
Looking to the previous habitat of larger quartz rock
Showing the small shale ridges
Pebbles and plants present
Plants absent
Viewing site from the north
Also looking SE towards Die Kop and Potberg
Looking SE towards Die Kop and Potberg
MBB7266 SE Lemoenpoort, view to southwest
13. A view from south Dwarsindieweg.
4c. View northwards from Bakoven
7. View south to Villiersdorp
6. Near east end with P marking the site of Haworthia pubescens, R indicating Rooiberg to the east
5. Looking north with M marking the location of Haworthia maculata, K – Karoo Garden
4. Still further east looking west northwest
3. Same as figure 1, but from further east
2. Same spot looking east northeast
1. Looking down the north face Die Nekkies east to west
1f. West Muiskraal
1e. Looking southwest to Garcia Pass from same spot as above
1d. Looking eastwards over Muiskraal
1c. View looking west from north Muiskraal
1b. View looking south west over Muiskraal to Garcia Pass
1a.View from above Onverwacht North Westward to Ladismith
Towerlands Eastward
Towerlands Westward
Towerlands
View from the upper slopes of the Zuurberg at Wilgerfontein, with Darlington Dam just visible in the distance.
Fig. 8a. Habitat of H. elizeae. Stormsvlei.
Fig. 8b. Habitat of H. elizeae. Stormsvlei.
03 . View west of south from Sandberg.
View 2 From Cilmor eastwards.
View 1 From Cilmor to the northeast.
Fig. 43. May’s Mine scene.
Fig. 4 View to the northwest across the flood plain.
Fig. 3 View northwest from a central position.
Fig. 2 Some indication of the topography of the south bank.
In a series of postings in the summer of 2019, Bruce Bayer discussed the taxonomy of Haworthia retusa and related H. turgida, H. pygmaea, H. mirabilis, H. magnifica, H. emelyae, H. mutica, H. groenwaldii, H. floribunda. Here are supplemental references.
H. retusa (L.) Duval in Pl. Succ. Hort. Alenc.: 7 (1809): Aloeretusa L. (1753). Lectotype, designated by Scott (1985): Illustration in Commelin, Horti Med. Amstelod. 2: t. 6 (1701). Epitype, designated by Breuer & Metzing (1997): Blikbonnie, E Riversdale, Dekenah s.n. NBG144772 (NBG).
retusa: with leaf-tips bent back thumb-like.
Rosette stemless, slowly or seldom proliferous, to 12cm φ. Leaves 10-15, turgid, rigid, with pronounced retused end-area, pointed tips, variously lined and windowed. Without surface spination and usually without spination on margins and keel. Colours brownish or green and seldom purpling. Inflorescence simple, robust, to 30cm. Flowers compacted on inflorescence, white with greenish-brown veins.
H. retusa var. turgida (Haw) Bayer (2012)H. turgida Haw. in Suppl. Pl. Succ.: 52 (1819). Neotype, designated by Breuer & Metzing (1997): Swellendam, Breede River Bridge, Bayer 2420 (NBG).
turgida: swollen.
Rosette partially stemless, proliferous, 5-10cm φ. Leaves 20-40, turgid often as thick as broad, recurved or slightly retused, generally mottled, yellow-green to pink in sun, margins and keel lightly spined. Inflorescence simple, 15-20cm. Flowers slender, brownish-white with darker venation.
H. pygmaea V.Poelln. in Repert. Spec. Nov. Regni Veg. 27: 132 (1930). Neotype, designated by Breuer & Metzing (1997): hills E Great Brak, Fourcade 4759 (BOL).
pygmaea: dwarf.
Rosette stemless, slowly proliferous, 6-10cm φ. Leaves 12-15, retused, round-tipped, surface pellucid with obscure raised tubercles, sometimes intensely papillose. Inflorescence simple, robust, to 30cm. Flowers white with greenish veins.
H. mirabilis (Haw.) Haw. in Syn. Pl. Succ.: 95 (1812): Aloemirabilis Haw. (1804). Neotype, designated by Bayer (1977): Illustration in Curtis’ Bot. Mag.: t. 1354 (1811). Epitype, designated by Breuer & Metzing (1997): Skuitsberg, between Caledon and Greyton, Bayer 2453 (NBG).
mirabilis: wonderful.
Rosette stemless, proliferous, to 7cm φ. Leaves 10-15, retused, 3-4cm X 1,5cm, markedly retused, acute above, face translucent and lined, dark green, with marginal spines turning reddish in the sun. Inflorescence slender. Flowers narrow, elongate, biarcuate bud, upper lobes pinched at tips.
H. mirabilis var. magnifica (V.Poelln.) Bayer (2012): Haworthiamagnifica V.Poelln. in Repert. Spec. Nov. Regni Veg. 33: 240 (1933): H. maraisii var. magnifica (V.Poelln.) Bayer : 131 (1976). Lectotype, designated by Breuer & Metzing (1997): Riversdale, Ferguson.
magnifica: magnificent.
Rosette stemless, slowly proliferous, to 8 cm φ. Leaves spreading, retused to ground level, dark green to purplish, scabrid to finely spined margins, end-area slightly translucent between the veins, surfaces with small slightly raised tubercles. Inflorescence slender, to 40cm. Flowers brownish veined, few open, upper lobes pinched at tips.
H. emelyae V.Poelln. in Repert. Spec. Nov. Regni. Veg. 42: 271 (1937). Lectotype, designated by Breuer & Metzing (1997): [unpublished image] (B).
emelyae: for Emely Ferguson.
Rosette to 10cm φ, seldom proliferous. Leaves 15-20, distinctly retused, pointed, barely translucent, with scattered elongate small flecks, with obscure raised tubercles, lined, reddish-brown hued. Inflorescence simple. Flowers 15-20, white.
H. mutica Haw. in Revis.:55 (1821). Lectotype, designated by Bayer (1978): [image] (K; later published in Excelsa 8: 50 (1978). Epitype, designated by Breuer & Metzing (1997): NE Soetrivier Bridge, Bayer KG623/69 (NBG).
mutica: without a point.
Rosette stemless. non-proliferous, 6-8cm φ. Leaves 12-15, retused, blunt-tipped, brownish-green, in habitat developing purplish cloudiness, barely pellucid with several longitudinal lines. Inflorescence simple, to 20cm. Flowers white with brownish veins.
Description: Always growing singly, but in a couple of cases growing in groups of plants of different ages. Rosette: acaulescent, 40-50 mm Ø (exceptionally up to 100 mm). Leaves: 5-7(8), fleshy, spreading, end-area in most cases very rounded (resembling H. springbokvlakensis), 20-25 mm long, 13-18 mm broad, 11-13 mm thick. Leaf surface of back and leaf-base, smooth, opaque; colour dull dark green. Margins: opaque, smooth. Back without keel. End-areas: 13-18 mm long, face translucent, with 3-5 whitish parallel lines, the middle one the longest (sometimes parallel lines are ± partly of greenish colour). Surface of the end area, rough with small papillae. End area of some specimens are with ± whitish, ± raised dots, in a few cases the dots are blended into cloudy flecks. Leaf-tip: very rounded, without teeth. Inflorescence: 300-600 mm long, 1.2-2 mm diam., Peduncle: 200-300 mm long. Sterile Bracts: 15-20, 5-7 mm long. Raceme: 100-300 mm long. 8-18 flowers, 2-5 flowers open at one time. Fertile Bracts: 3-4 mm long. Pedicels: 2-4 mm long, 1-1.2 mm diam. Flowers: 14-16 mm long. Perianth: 3 -4 mm diam. Flower-face: 9-12 mm long. Upper part: 6-7 mm long, 3-5 mm broad. Lower part: 5-7 mm long, 5-9 mm broad. Fruit: 11-18 mm long, 2.5-4 mm thick. Flowering time February–March. Type Information Collector: J. Groenewald s.n. (= MBB7801) Locality: 3420 (-BA), Buffeljagsrivier, Western Cape, SA.
Haworthia groenewaldii ‘typical’ specimen with whitish flecks and parallel lines with some green colour.
Haworthia groenewaldii in Habitat. Specimen without whitish flecks, growing in between Merxmuellera disticha, a hard and unpalatable grass.
Another specimen of Haworthia groenwaldii with little whitish flecks
A specimen of Haworthia groenewaldii with inflorescence among a group of smaller plants.
H. floribunda V.Poelln. in Repert. Spec. Nov. Regni Veg. 40: 149 (1936). Neotype, designated by Bayer (1982): [unpublished image] (B). Epitype, designated by Breuer & Metzing (1997): Blackdown, NE Heidelberg, Bayer 158 (NBG).
floribunda: many flowered.
Rosette stemless, up to 3cm φ, slowly proliferous. Leaves 20-30 dark green opaque, upto ovate-lanceolate, spreading, twisted with flattened, rounded tip, margins scabrid to dentate. Inflorescence simple, to 250mm. Flowers 10-15, greenish-white, few open together.
Journal of the Linnean Society vol 18 – 1881, J. G. Baker ‘On Aloinea and Yuccoidea’ – 3. Haworthia starting on 209 (page 197). H. retusa 220 (208). H. turgida on the next page.
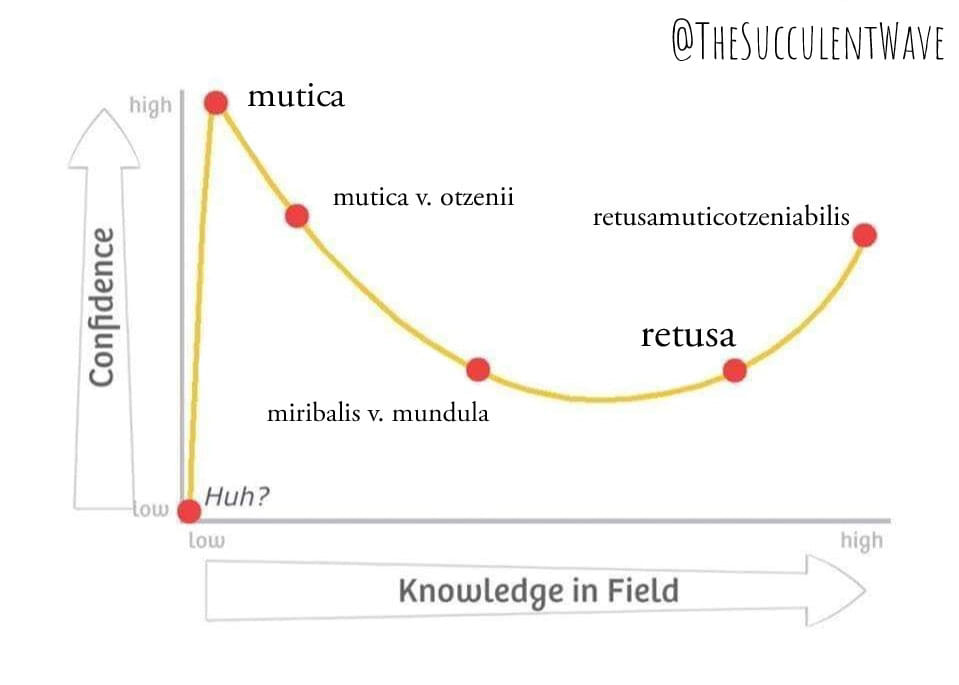
Everything looks easy to those who haven’t a clue.
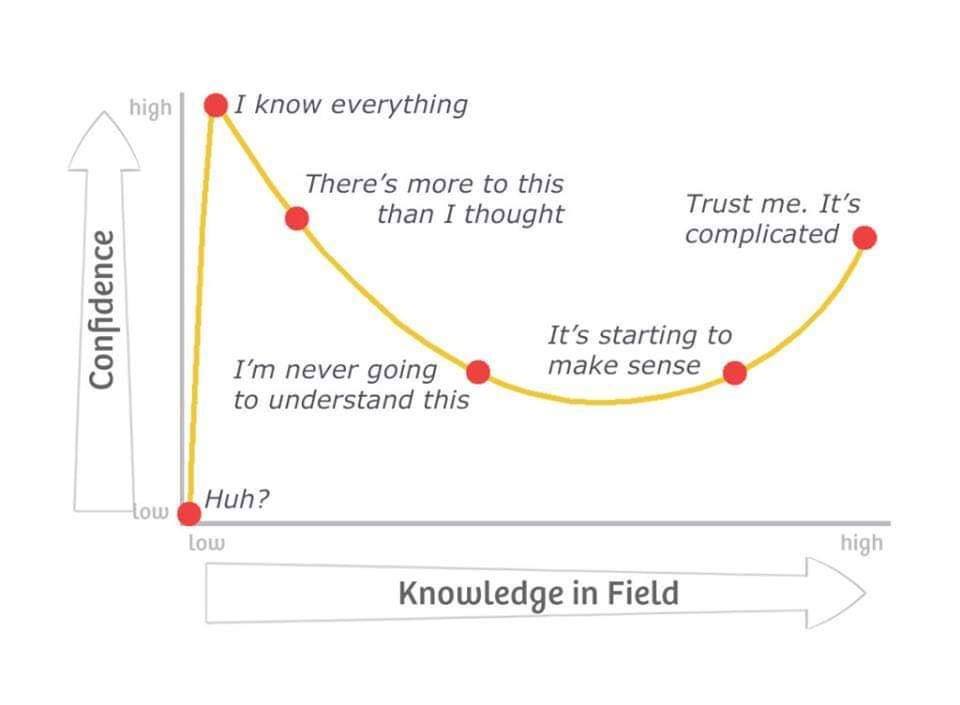
In other words – the Dunning-Kruger Effect is essentially that we who do not know enough to recognize our incompetence? This could not be more true of the fields of science and religion. Lawrence sent me some great stuff on taxonomy that demonstrates this too well with regard to both Darwinism and the Linnaean system.😅
I would like to add to that confidence/knowledge field because at the point of “never understanding”, the graph branches. One direction is intellectual superiority (no one else can understand this better than me), the other is … how do I get myself and others to grasp that we are all equally ignorant and that knowledge of the material world has no reality.
I really like that post of Steven’s re Dunning-Kruger Effect because it supports my contention that unless we recognize what we will not i.e. this is a conscious creation, we will never understand anything!
Very important. Naming species after people is in fact very suppressive as questioning the validity of the name gets confounded with the persona. Steven Hammer is worthy of considerably more than this absurd non-entity that I doubt he would have been party to. The same applies to H. bobbii that has an identical history. Breueri, jakubii, even bayeri has its worms as Gerhard is anxious to inform us all. So does marxii where a shepherd who so remarkably brought it to the attention of mankind via a farmers wife. Kobus Venter and then myself get an amazing acknowledgement from the author of the name. How could it have been undiscovered for so long?
(ed. – original Dunning Kruger paper – Unskilled and Unaware of it: how difficulties in recognizing one’s own incompetence lead to inflated self-assessments.)
(ed. – try internet searches for “non-linnaean taxonomy”, “Raymond Hoser problem”.)
Hi Bruce, I would like to hear your opinion on the flowering time of H.magnifica var splendens, Haworthia truncata and some other species whilst it is very dry at the habitats where they grow. (I include two photos of the species mentioned) Without rain, the plants have lost their condition,yet they use water and energy to produce their flowers. On top of that, other insects and animals have limited choice to their feeding menu and would gladly include a haworthia flower stalk. I keep in mind that the survival of a plant at a particular site depends on the existence and activities of pollinators. Could it be that these plants adapted to the availability of pollinators in summer and have found a way to preserve enough moisture to flower during the dry months of the year?
Bruce Bayer What a nice surprise Emile. Grower extraordinaire! Splendens flowers like all mirabilis (?) in February. Retusa (flowers October), emelyae in the Karoo flowers October. I think truncata also flowers summer, but really irrelevant here. In my current ideas is that splendens (mirabilis), retusa (turgida, pygmaea) and emelyae are in reality, one true basic life form i.e. species. Rain is not a significant (note – significant) factor in flowering time.
Emile Heunis Thank you for the complement. I have had you as a mentor and inspiration,visiting Karoo Botanical Gardens in the late 1970’s and early 80’s. I bought many beautifully grown Haworthias when you had sales at KBG in the 80’s. I use the names as I have come to know them. Please appreciate that I have no understanding of the species concept in Haworthia or any plant form. I have always avoided any comment on the topic because of that. As an artist with a life long interest in nature, particularly succulents, my interest lies in their shape,form and colour, qualities that allow them to survive in nature. As most human thinking support (sadly), all that is to please Homo, the one that can determine the existence of all other life forms. My love for nature brings many questions. The magnitude,the diversity and how it exist together in peace and at war with one another. Hence my question. How did it come that ,in this case, H. splendens survived on a particular spot whilst that spot is not ‘ideal for producing flowers. As it is, it is ideal!! The plants are in excellent condition, at least the two times I could go there, and as can be seen in most photos taken by other enthusiasts. Could you please explain your concept of ‘significant factors’ that do play a role?
Bruce Bayer Thank so much Emile. Funny I saw “splendens” originally a short way from the now fenced spot. They were larger and more colorful in a sandier environment. While we have a mechanistic concept of species and their environment we will never understand either of these. A species is a fractal system based on metaphysical realities with infinite possibilities. The world too is a “living” thing. There are five main energy systems (elements) in creation related to the five regular figures that can be fitted to a sphere – hence the “tree of life”. Plants have one of these, insects two, snakes reptiles birds, three, mammals four and man at top of creation five. Always infinite possibilities with the energy fields starting at math zero or infinite zero. There is far more to creation that our absurd “science” belief system allows.
Emile as an extra. I have seen out of season flowers on both mirabilis and retusa and what is more have encountered field hybrids of these as well as of these with floribunda. The complexity of the interaction between all these so-called “species” is incredible.
ed. ~ original correspondence on Facebook 27 January 2021.