Written for, and then printed and distributed by Haworthia Study Group, New South Wales – October 1970.
M. B. Bayer, Karoo Garden, Worcester, South Africa.
Much of the confusion in the taxonomy of the genus Haworthia could have been avoided if more attention had been paid to localities and inherent variability of the species. The tendency has been to base species on single specimens and to regard the species as variable concepts subject to personal opinions. According to Stebbins in his book “Variation and Evolution in Plants”, a species comprises a system of populations separated from each other by complete or sharp discontinuities in their variation patterns, and this must have a genetic basis. Therefore there must be isolating mechanisms to prevent transfer of genes. In Haworthia, most (not all) of the species are found in rock formations and stable situations at moderate altitudes, and hence the populations are well spaced. Haworthias are also insect pollinated, mostly by solitary bees, and as stated by Clausen in “The Evolution of Plant Species”, it is logical that spatial separation of the plant populations coupled with flight limitations of the pollen vectors, will with time have led to differentiation of highly localised populations. A further problem in Haworthia is that the species do in fact exist at different stages of differentiation. Thus one may be forced for practical reasons to regard a widely ranging series of varying populations as a single species, simply because the degree of inter-gradation and variability precludes any other alternative. If there is an alternative, it is to recognise varieties in the sense that this concept has been used in the past, i.e., to apply to individual variants and forms.
47. Haworthia glauca Baker, JLinn. Soc. 18:203(1880). V.Poelln., Cact.Succ.J(U.S.) 5:30(1936). ibid., Feddes Repert.Spec.Nov. 43:108(1938). Bayer :118(1976). Bayer :67(1982). Scott :49(1985). Type: Cape, Zuurberg Pass, Cooper (K): H. carrissoi Res., Bolm.Soc.Broteriana 15:161(1941). Type: Cape, Zuurberg Pass. Not preserved.
glauca: glaucous, powdery, bluish-green.
Rosette to 80mm φ, caulescent, proliferating. Leaves many, to 60 X 15mm, erect spreading or incurved, scabrid, glaucous grey-green, with or without tubercles. Inflorescence simple or occasionally compound, to 300mm. Flowers tepals fused, tube straight. Lower inner tepals revolute.
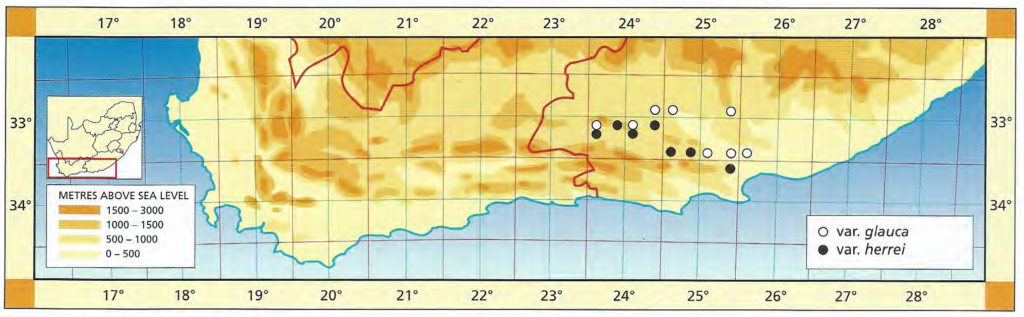
1982 – Baker gave the locality for H. glauca as ‘Orange Free State’ which is clearly wrong. The actual locality is probably in the Zuurberg mountains north of Uitenhage. This is however, only the eastern range of the species and it is not known further east than the Zuurberg Pass itself. Up here on the mountains in sour grassveld, the plants are very much like described by Baker. The leaves are glaucous, glabrous and incurved, while the stems are not unduly elongate. The full distribution of the typical species is not known. From Paardepoort in the Klein Winterhoek mountains and westwards, the species is much more variable. The stems of the plants are usually well‑developed, the leaves may be erect or spreading and are tuberculate even if only slightly so. This is the basis for recognising the var. herrei. H. glauca var. herrei fa armstrongii comprises a localised population northeast of Uitenhage ‑ based on a vegetatively propagating clone. J.R. Brown said that this form was close to ‘ H. herrei var. depauperata’ in general appearance and colour, but easily separated on the basis of having non‑confluent tubercles only on the margins and keel. The one distinguishing feature of fa armstrongii is colour ‑ both in habitat and in cultivation it tends to be darkish green and lacks the glaucous sheen of the typical species. The fa armstrongii can be superficially compared with H. fasciata except that it is caulescent. It also occurs in close proximity to H. fasciata fa browniana. Schneider (1972) showed that fa armstrongii and H. glauca were both distinguished by the absence of waxing in the stomatal chambers, and this too suggests that they belong together. In a very recent publication, Brandham and Cutler (1981), show that the fa armstrongii is a pentaploid of probable hybrid origin, showing closer affinities to H. coarctata than to H. glauca.
1999 – There is a small discrepancy in the citation of the type as Col Scott cites ‘Zuurberg Pass, Cooper sn.’ whereas Breuer and Metzing state ‘Cape without locality’. Although it may be supposed that the var. herrei is a discrete element, it is not possible to separate the herbarium specimens in this way. The var. glauca comprises the smooth eastern form, but this also occurs even west of the range for the more commonly tubercled var. herrei. The range of variation in that variety is quite considerable, as is apparent from the many species and varieties which emanated from particularly the Steytlerville area.
a. var. glauca
Distribution: 3224 (Jansenville): Fairview (-CD), Bayer 4179 (NBG); 1km N. Jansenville (-DC), Branch 321 (NBG). 3225(Somerset East): De Hoogte (-CD), Smith 3521, 5547 (NBG). 3323(Willowmore): Wilds View (-BA), Vlok 1554 (NBG). 3324(Steytlerville): Burbank (-AA), Marx 58 (NBG); 2km E. Humefield (-AA), Bayer 4168b (NBG). 3325(Port Elizabeth): Paardepoort (-AC), Archibald in NBG1320/37 (NBG), Bayer & Venter 6615 (NBG); De Plaat (-AD), Venter 91/16 (NBG); Zuurberg Pass (‑BC), Smith 12, 1183 (NBG), Marloth 4212 (PRE); Zuurberg Pass, N. Inn (-BC), Archibald 5280 (PRE).
Inadequately located: Somerset East, Smith 1184 (NBG); ex hort Ross-Frames in NBG75/44 (NBG), Britten 5748 (NBG), Henderson 1442 (NBG).
Haworthia glauca var. glauca JDV91/16 northeast of Kirkwood. It occurs here with tuberculate specimens.
Haworthia glauca var. glauca JDV90/12 Zuurgerg Pass. Without tubercles and with incurving leaves.
b. var. herrei (V.Poelln.) Bayer :122(1976). Bayer :118(1982). H. herrei V.Poelln., Feddes Repert.Spec.Nov. 26:24(1929). ibid., Cact.Succ.J(U.S.) 1930. Type: Cape, STE 5682. Not preserved. Neotype (B&M): Campherspoort, Barker 5069 (NBG): H. herrei var. depauperata V.Poelln., Feddes Repert.Spec.Nov. 31:86(1932). Type: Sundays River, between Steytlerville and Port Elizabeth, STE 6642. Not preserved: H. jacobseniana V.Poelln., Desert Pl.Life 9:102(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1038). Type: Cape, Jansenville, Mrs E. Jones in Triebn.944. Not preserved: H. eilyae V.Poelln., Kakteenkunde lO:152(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1938). Type: Cape, Steytlerville Distr., Archibald 1125. Not preserved: H. jonesiae V.Poelln., Kakteenkunde lO:152(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1938). Type: Cape, Steytlerville, Mrs E. Jones. Not preserved: H. herrei var. poellnitzii Res., Mems.Soc.Broteriana: Succ.Afr. 3:89(1943). Type: Cape. Not preserved: H. eilyae var. poellnitziana ibidem. Type: Cape. Not preserved: H. eilyae var. zantneriana idem. :90 loc.cit. Type: Cape, STE6625. Not preserved. H. armstrongii V.Poelln., Kakteenkunde 10:152(1937). ibid., Feddes Rep.Spec.Nov. 43:108(1938). Jacobsen 2:537(1954). H. glauca var. herrei fa armstrongii (V.Poelln.) Bayer :99(1976). Bayer :99(1982). Type: Cape, Adelaide, Armstrong. Not preserved. Lectotype (B&M): icon (B).
herrei: for Dr.H. Herre.
None of the species or varieties described here are represented by type specimens nor good locality data, and it is perhaps fortunate that it is not very important in this case. I wrote an article for the New South Wales Study Group in 1970 which included photographs of plants collected from the Campherspoort vicinity. The variation in the clones covered easily the characteristics attributed to the elements described: narrow stems with erect incurving leaves, wider stems with erect spreading leaves, short nearly deltoid leaves, long lanceolate leaves, tubercled or non-tubercled. There has been some cytological evidence to suggest what relationship the fa armstrongii has to the species. However, the results are conflicting and it is not easy to question this (or scanning electroscopy for that matter). Brandham and Cutler (1981) report H. glauca to be ‘mostly hexaploid….but two .. were aneuploids’. Their discussion is ambivalent as on p515 of the article they suggest there is insufficient evidence to decide its parentage, whereas on p540 they state that it cannot be a variety of H. glauca because it is pentaploid and the species is hexaploid. The authors refer to two populations of H. glauca studied, but the list of materials cites only one (actually the var. herrei), and one for the fa armstrongii. Riley (1959) reported H. glauca to be tetraploid and thus a pentaploid within the species is to be expected. Finally it is now known that H. glauca is common in the area close to where fa armstrongii was originally collected. A recent visit to the site suggests that it no longer occurs there. Brandham and Cutler also looked at epidermal character where they conclude that fa armstrongii was closer to H. coarctata than to the Steytlerville collection of H. glauca. This result has to be viewed in proper perspective. It is an hypothesis based on a limited range of material and it conflicts with Schneider’s results (quoted above). Even the casual observer could probably conclude that the most logical complement to H. coarctata, both geographically and morphologically, is H. glauca. My own experience of ESM studies (and similar sophisticated techniques) is that not enough is done to first examine general variation, and that conclusions only apply within the narrow limits of the material studied.
Distribution: 3323 (Willowmore): Redcliffe (-BA), Bruyns 7053 (BOL); Miller (-BB), Acocks 19902 (PRE); 4km SW. Miller (-BB), Smith 3658 (NBG); 11km SW. Miller (-BB), Smith 3660; W. Knoetze (-BB), Smith 6917 (NBG); 11km Knoetze to Miller (-BB), Smith 6863 (NBG). 3324(Steytlerville): Campherpoort (-AA), Smith 7046, 7047, 7048, 7049, 7050, 7055, 7056, 7058, 7059 (NBG); 1km W. Campherpoort (-AA), Smith 3648, 3648a (NBG); 6km W. Campherpoort (-AA), Smith 3652 (NBG); 16km W. Campherpoort (-AA), Smith 3237, 3655, 3655a Pienaarspoort (-AB), Smith 6217, 6866 (NBG); E. Campherpoort (-AB), Smith 3592 (NBG); Waaipoort (-AD), Bayer & Bruyns 6584 (NBG); Zeekoeisnek (-BC), Bayer & Bruyns 6586 (NBG); Kleinspoort (-BD), Otzen in NBG618/41 (NBG), Smith 5170 (NBG); 3km S. Kleinpoort (BD), Smith 2924, 2924a, 2924b (NBG). 3325(Port Elizabeth): Bauerskraal (-CB), Bayer & Venter 6598 (NBG); 10km NW. Uitenhage (-CB), Smith 706, 3076, 7195 (NBG), Smith 912 (NBG,PRE).
Inadequately located: Kirkwood to Jansenville, Cunningham & Cook in NBG761/31 (BOL); Steytlerville, Taute in NBG950/37, Smith 2754; Pillans in NBG356/16, Peers in NBG68046, Malherbe in NBG489/42.
Haworthia glauca var. herrei JDV91/16 north of Kirkwood. It is the tuberculate forms which are predominant and several varieties were described from this polymorphous species.
Haworthia glauca var. herrei JDV91/121 Kleinpoort. Although supposed to have spreading leaves it does not appear as though even the geographic criterion for two varieties can be justified.
Written for, and then printed and distributed by Haworthia Study Group, New South Wales – October 1970.
M. B. Bayer, Karoo Garden, Worcester, South Africa.
Much of the confusion in the taxonomy of the genus Haworthia could have been avoided if more attention had been paid to localities and inherent variability of the species. The tendency has been to base species on single specimens and to regard the species as variable concepts subject to personal opinions. According to Stebbins in his book “Variation and Evolution in Plants”, a species comprises a system of populations separated from each other by complete or sharp discontinuities in their variation patterns, and this must have a genetic basis. Therefore there must be isolating mechanisms to prevent transfer of genes. In Haworthia, most (not all) of the species are found in rock formations and stable situations at moderate altitudes, and hence the populations are well spaced. Haworthias are also insect pollinated, mostly by solitary bees, and as stated by Clausen in “The Evolution of Plant Species”, it is logical that spatial separation of the plant populations coupled with flight limitations of the pollen vectors, will with time have led to differentiation of highly localised populations. A further problem in Haworthia is that the species do in fact exist at different stages of differentiation. Thus one may be forced for practical reasons to regard a widely ranging series of varying populations as a single species, simply because the degree of inter-gradation and variability precludes any other alternative. If there is an alternative, it is to recognise varieties in the sense that this concept has been used in the past, i.e., to apply to individual variants and forms.
Kaboega is located in the Zuurberg Mountains north of Port Elizabeth in the Eastern Cape province of South Africa. Read more about this in Haworthia Update Volume 1, Chapter 5:- The Haworthias of Kaboega. There are a mind-boggling array of Haworthia populations here in an area considered to be the meeting point of several vegetation biomes. There is much exposed rock, and the soil is very skeletal, composed of three major groups: sandstone, mudstone, and glacial deposits. These pictures are of a Haworthia cooperi variant that occurs high up on sandstone. I went to this spot because researchers had sent me a picture of a cycad festooned with Haworthia. I did not get to the exact spot but have seen the way it forms hanging bundles in other situations.
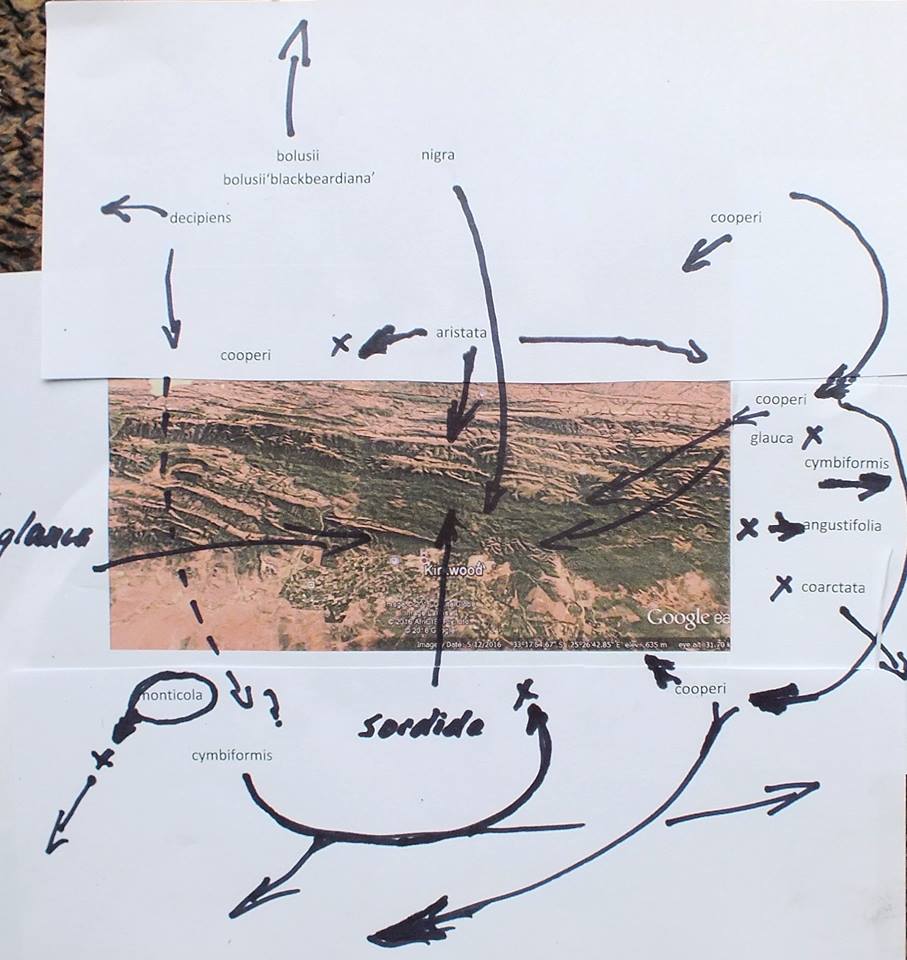
Haworthia glauca!! can also be found here. On Kaboega these plants often have a very close resemblance to H. coarctata and it is no co-incidence that the distributions of these two species complement each other. An essential element of species recognition is their juxtaposition and if they occur in very close association or not. Darwin said as much.
I visited four populations of this greenish cooperi. One can find plants like this from east of Grahamstown right through to the Little Karoo. Here they are on Dwyka (glacial) skeletal soil.
These next are in the shales low down in the valley on Kaboega – I name it H. aristata. It is very common in the area but complements H. cooperi while there are populations that are neither. Populations cannot be treated in isolation and there is a distinct possibility/probability that I have been too generous with species. The attempts to find answers via DNA sequencing should make the vendors of that technology thoroughly ashamed.
More of these green things. I would guess that these would class as the simple progenitors of cymbiformis and cooperi. Perhaps even of mucronata?
This is Haworthiopsis sordida that does not occur, as far as I know, north of this. H. nigra also occurs here at it’s most southern at this longitude. Altogether it is quite a complex network of distribution patterns that relate to greater plant geography.
From another population as variants on a theme. have seen about 30 such just on this small mountain area and it just suggests what is still unseen on the length and breadth.
Not a great diagram but a way to appreciate the drammatic choreography of plant distribution and how it impacts on classification. Without it Haworthia names make no sense other than as imagined and fantasized. Cooperi and cymbiformis occur as intertwined species to the east and south. In the south they extend westwards to get lost in H. mucronata. Cymbiformis as an independent species does not enter Kaboega except as an observable variant of H. cooperi. The cooperi gets lost westwards as variants of H. decipiens. Perhaps close northwards as H. aristata. H. glauca does cross the Zuurberg but is here confused with H. coarctata that may occur in recognosable form on the eastern tip. Angustifolia is on the eastern end too but does not enter Kaboega. Neither do H. monticola or H. zantneriana from the west. This is also closely tied to the intrigue of winter vs summer rainfall and still further to the massive geological changes of the very recent..