
In a very interesting book by Stephen Gould entitled “Rock of Ages”, in which he propounds his principle of NOMA – non-overlapping magisteria. This states that science and religion should not be confused nor mixed.
So this is not a confession of confusion – you do not confess to what is obvious. It is an admission, and an admission can be construed as an apology. But, as a rhetorical question, how can one apologize and expect forgiveness when one continues to walk the errant path?
I started to write about Haworthia to dispel confusion, and yet more than 40 years on, this confusion has not become any less. The conclusion I have come to (and I wish it was a closure) is that the prime source of confusion is simply the human condition. In mystic philosophy one can read… “Born in ignorance, we live in ignorance and we die in ignorance”.
I think that my interest in Haworthia stems from my conscious effort to dispel this primal confusion and find some of the order in my view of creation. The classification of plants suggested just one small piece of my world which was available to me, and Haworthia as one group which no one else could explain to me. What have I now learned and what contribution does this make to dispel confusion?
My courage to now say something more directly arises from a recent request by SANBI to write a synopsis of Haworthia for an E. Cape Flora. I feel that I have done that fairly successfully. The problem is now to produce a similar product for the SW Cape and this is considerably more difficult.
To explain Haworthia one has to first cross the hurdle of the present classification of the Asphodelaceae and the perceptions that Aloe, Haworthia and other genera are clear discrete groups that the classification suggests. Ignore completely the issue of whether the so-called species are clearly separable. Haworthia consists of three quite discrete groups which have no closer relation to each other than do any other of the extant Asphodeloid genera. They each have their own problems and idiosyncrasies but it is the subgenus Haworthia which concerns me most.
In 1975 I prepared a map to illustrate the relationship of the SW Cape species (primarily south of the Langeberg Mts.) and this was published in Excelsa (5, 1975). In this map I tried to illustrate the problem of continuity which simply makes it impossible to recognize clear closed groups of plants which we can with any truthfulness say are species. It is the reality for Haworthia, and my conviction is that the model which underlies the Latin binomial system is fundamentally flawed. It works because our ignorance clouds its shortcomings.
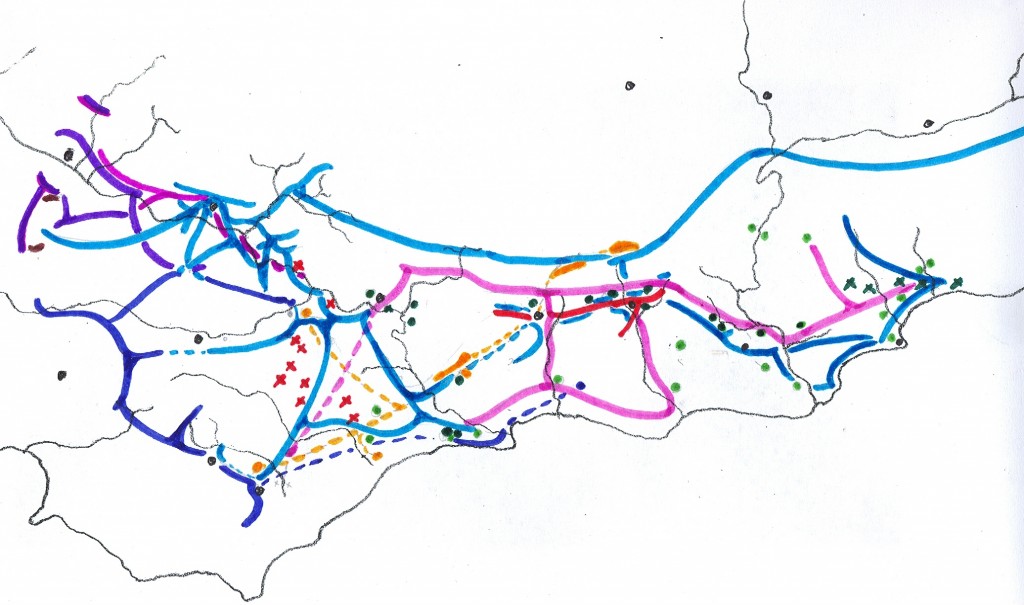
Map 1 Distribution of SW Cape species of Haworthia
Updating my map may show why (Map.1). It is perhaps possible to identify three main elements that are nevertheless extremely difficult to map. Very seldom do we find all three growing in close proximity and never do we find any of the components I suggest doing so either. We often struggle to suggest which of the three we are being faced with.
The names we can use for these three are:-
H. retusa – to include or cover mutica, turgida and pygmaea
H. mirabilis – to include or cover maraisii, magnifica, emelyae and heidelbergensis
H. floribunda – to include or cover parksiana, chloracantha, floribunda and variegata.
There are other elements viz:-
H. herbacea – to include or cover reticulata, maculata and pubescens
H. rossouwii –
But this is a simplification and not a solution.
What simply has to be recognized as a fact of life that this problem is not of my creation and I do not believe that the use of any technical sophistry, trick of classification or nomenclature will possibly dispel it. These “species” are as interconnected as the strands of a complex web as intricate as any a confused spider could construct.
Essentially one has a mid- to late-summer flowering group and a spring flowering group, but they are linked. H. pygmaea, in the retusa complex, actually springs from the mirabilis complex; while its counterpart mutica in the west, seems to spring from the retusa complex. Emelyae, in the mirabilis complex, seems to be linked back to rossouwii via multifolia. These are the kinds of realities one has to deal with.
The relationship of the three main elements I have mentioned differs from point to point on the map, and to describe these relationships means that one has to virtually deal with populations and groups of populations one-by-one. I have done that, particularly in the book Haworthia Update Vol. 1n and again in Vol. 2. But since the drafting of the last manuscript, I have been to the field again and found more material, which I will show now.
Essentially there are three areas which I will cover:- a. the southern Potberg and where the Breede River enters the sea south of Swellendam (Map 2); b. secondly Klipfontein at the northwest of the Potberg (Map 3); c. thirdly an area just west of that which I also call Die Kop, Wydgelee (Map 4). The localities are not indicated precisely on these maps but the detail is in archived and available information.
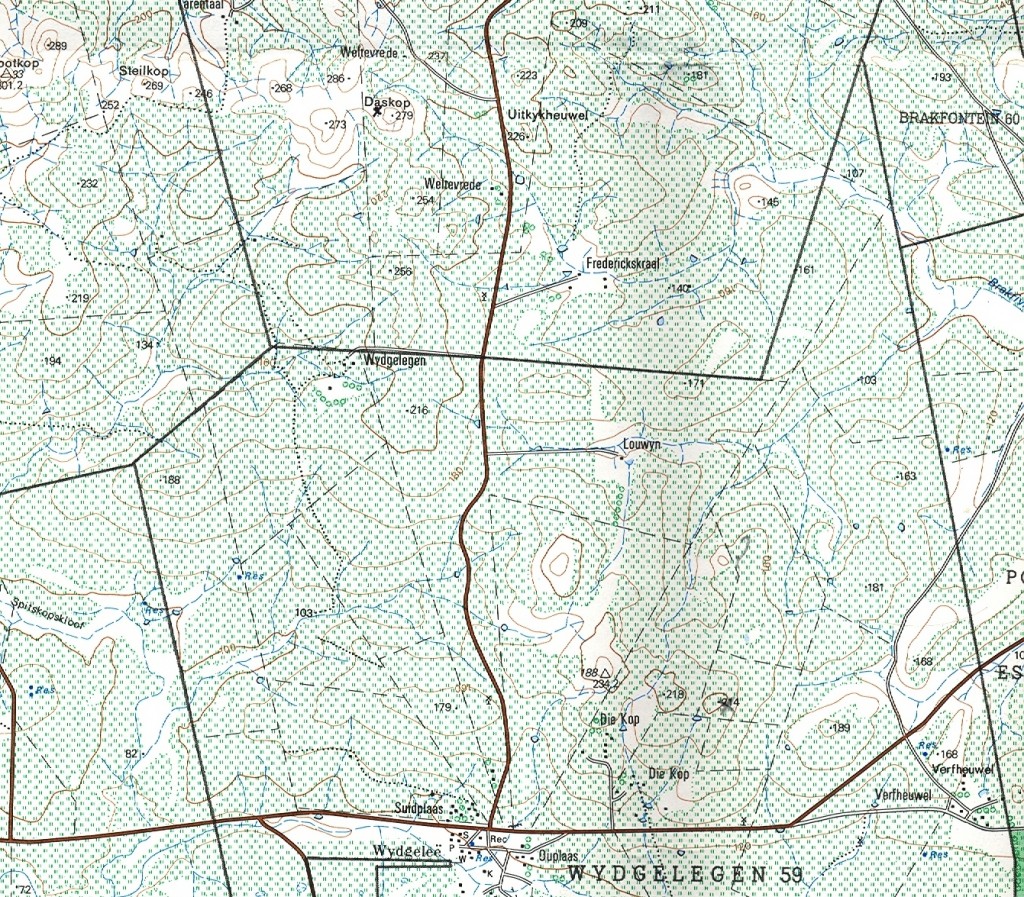
Map 3. Die Kop, Wydgelee. 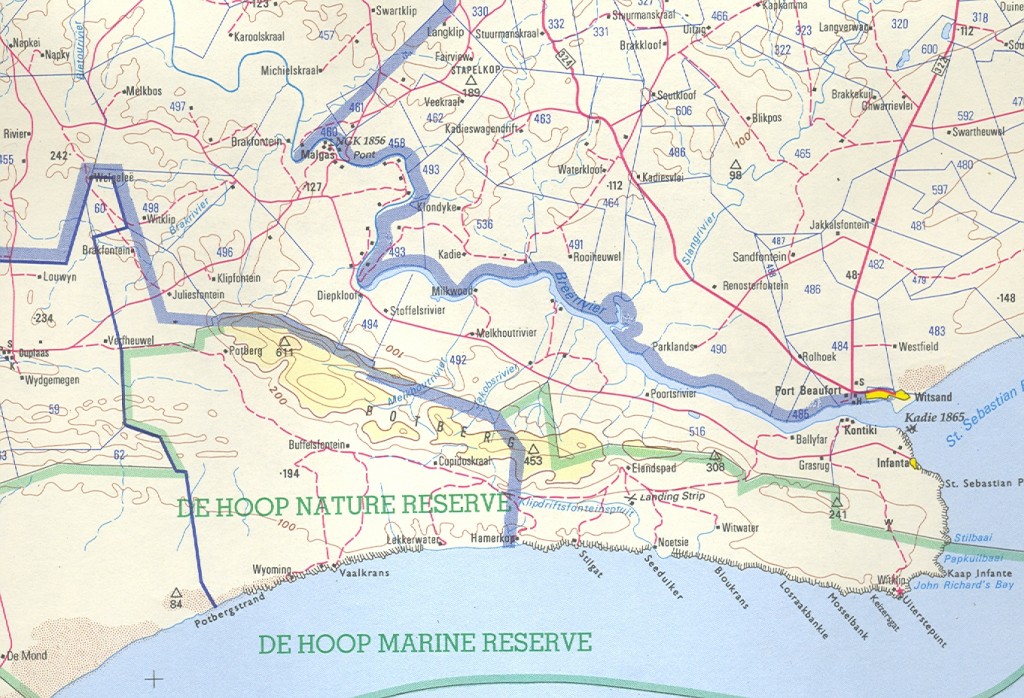
Map 1. Southern Potberg 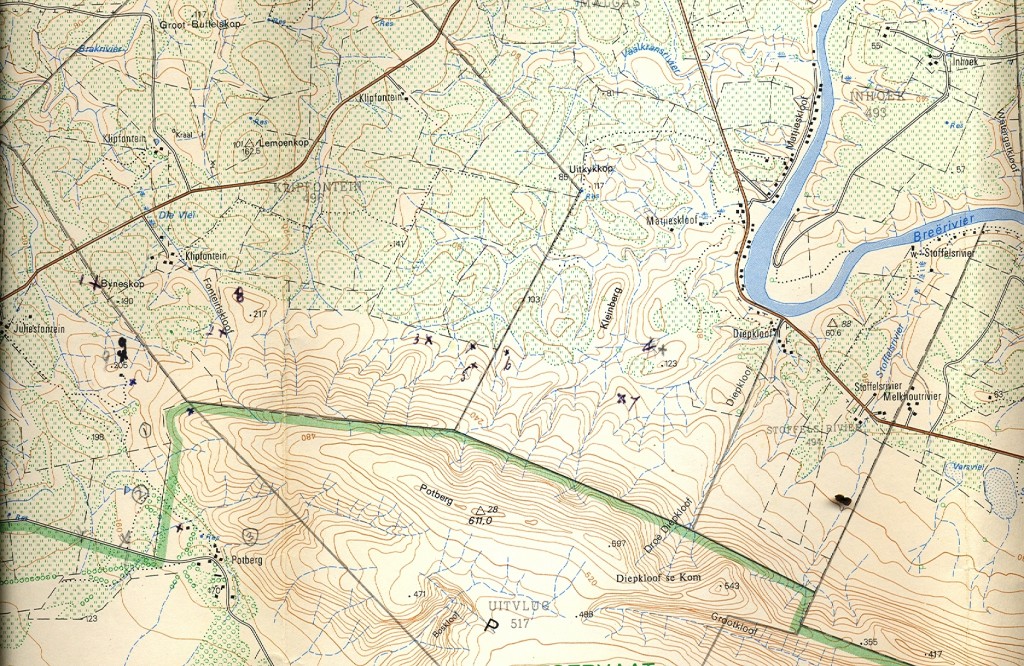
Map 2. Klipfontein, Potberg.
Results:
a. The southern Potberg.
Two of three relevant populations are discussed in Chapter 15. These are ADH594 Sandhoogte, and MBB7248 Lower Breede River H. mirabilis (pilosa). I refer to CB2018 and I can now report and illustrate it myself. This is MBB7251 (Figs 2a & b) from Buffelsfontein within the DeHoop Nature reserve. In the field the plants are not distinguishable from ADH 594 and grow in exactly the same Fynbos habitat viz a very rocky and grassy streamside slope with a dry northern aspect. The illustrations are of plants cultivated and hence far greener and softer than in habitat. What is striking is the tendency to surface spination which is such a feature of MBB7248.








b. Northwestern Potberg.
Some of these populations are also covered in Chapter 15. These are MBB6890, MBB6886, MBB6882, MBB6889 and MBB6544 (figs A18…22). My comment that H. variegata occurs here too should be borne in mind. This is on the upper slopes of the extreme western end of the Potberg. There is another record of C Burger’s from further east and south of Diepkloof for which I coined the name H. variegata var hemicrypta. Some doubt will now be thrown on the wisdom of that decision. The mirabilis populations referred to above are on the farm Juliesfontein and my new records are now east of that towards Diepkloof along the Breede River.
1. MBB7487 H. floridunda. Byeneskop (figs 3a to g). No good opinion is ventured in Chapter 15 concerning the populations noted above. Those populations seem to fall in the context of an interaction between H. maraisii/heidelbergensis/floribunda and I warn of infusion of H. variegata. The Byeneskop is on the very eastern border of the farm Juliesfontein and the plants are surprisingly and confidently comparable with the typical form of H. floribunda. The leaves are generally fairly strap like and flattened to the characteristic twisted ends as evidenced in floribunda. A few plants (see fig.3f) do have shorter and stubbier suberect leaves which characterize so many collections that confound the recognition of the relevant species. This particular habitat can be described as Renosterveld on silcrete with dominance of white clay and much grass and other succulents. Similar plants were also seen about a kilometer eastwards on the verge of vegetation transitional to Fynbos.







2. MBB7492 Klipfontein East (figs 4a to d). This is again a typically H. floribunda-like population. Whereas the preceding tends to be slightly tuberculate with the rougher greyer texture and colour of H. maraisii which would suggest floribunda var dentata, these plants tend to have a smoother leaf surface. Pictures of the flower are included although of very little diagnostic significance. The habitat is a steep rocky sandstone hillside vegetated with low Fynbos species.



3. MBB7494 Klipfontein East (figs 5a to d). The plants suggest H. floribunda except that the colour of the leaves is greener and there is a tendency to translucence, or less opacity of the leaves. The white flecks in the leaves are quite conspicuous and are more often encountered in H. turgida. Their significance taxonomically is probably nil as these markings/inclusions may appear in plants as remote as H. angustifolia var baylissii on the Zuurberg, E. Cape. The habitat is again short grassy Fynbos but unusual because of the presence of finely textured white quartz rock.



4. MBB7495 Klipfontein East (figs 6a to c). While we have been looking at plants on a series of buttresses of the Potberg, these plants are now closely associated with large white quartz boulders. They are small and although very similar to the preceding, the leaves seem to have lost the characteristic rounded ends and the margins are relatively strongly toothed.



5. MBB7496 Kleinberg (figs.7a to g). This population is only about 1km east of the preceding but there is a transition to Renosterveld. The habitat is again silcretious with the typical white clay substrate. This seems to be a very erodable surface and plant cover is quite low. The plants are surprising because they depart quite radically from the floribunda-like mode of the preceding populations. They are now distinctly magnifica-like and particularly reminiscent of var atrofusca except much smaller. Curiously a population across a small valley southwards in sandstone and Fynbos is in keeping with the floribunda-like finds westwards.







6. MBB7499 Juliesfontein (figs 8a & b). We returned to Juliesfontein to look at a promising rocky slope between Byeneskop and our previous closest collection on Juliesfontein. These are almost directly north of H. variegata and as can be seen from the pictures they revert to the forms described in Chapter 15 for that area viz. there is a similarity to heidelbergensis. The more leaves are more frequently pointed rather than rounded and translucence and venation is more conspicuous.


c. Die Kop, Wydgelee.
The nearest plants dealt with in other chapters are MBB7539 at Tarentaal that I liken to magnifica var atrofusca and MBB6030 at Napky which is maraisoid with floribunda-like character (type locality for H. maraisii var simplicior with rotate leaf-tips). It is important to note that the Tarentaal plants are not uniformly larger that my identification suggests and also that H. mutica is also present at its most southeasterly locality known south of Napky. Tarentaal only just appears on the map and the mutica locality is just off the map to the upper right (northeast) corner. It should be noted that populations assigned to H. heidelbergensis occur to the west at Kathoek, Beyersdal, N. Windheuwel and terminating in the contentious var minor at Rooivlei north of Bredasdorp. I think if an overview was now made of all these populations I have assigned to heidelbergensis in different chapters and in different publications, it will have to be concluded that it may represent the pivotal element in the whole classification process. Where it was the least represented species in the period preceding publication of Haworthia Revisited it is now far better known and any hypothesis prepared to test relationships in these southwestern Cape species Haworthia will need to question this centrality. Die Kop is a set of 5-6 silcrete and ferricrete hilltops of varying size and height but otherwise not significantly different from a large number of such habitats extending from Stormsvlei in the northwest and terminating against the Potberg and Breede River in the southeast. Collections from several of these have already been reported on in Chapter 15 with the general observation that their identification is in the uncertain linking maraisii and mirabilis. I hope, in the minds of readers, that I have not excluded heidelbergensis from this milieu.
1. MBB7500 Die Kop East (figs 9a to v). I doubt if any collection has generated such reservations in my mind as this one has and many illustrations are used to express the problem. The plants are large and up to 80-100mm in diameter. They are closely spaced in a small grassy, non-rocky area which seems to be geomorphically stable. They are extremely variable and resemblances extend from badia to retusa to mutica. The leaves may be short and rounded or fairly long and attenuate, closely adpressed to the ground (recurved) or sub-erect. One notable feature is that the plants have a milky colouration to the leaves that Seven Hammer once remarked on as a feature of H. mutica. While variability is known to be great, it is seldom that it is as dramatic as this in a population of plants as they occur in the field.





















2. MBB7502 Die Kop West (figs 10a to c). The plants here are smaller and more widely distributed and scattered over a larger area. The habitat is skeletal and rocky and the vegetation more herbaceous than at the eastern site. The similarity of the plants to those of 7500 is close but the variation is less extreme. There is less evidence of a mutica influence and a return to more or less the maraisii/floribunda/heidelbergensis archetype.



3. MBB7503 Die Kop Mid (figs. 11a to i). The first impression I had was that the plants included more mirabiloid influence. Not as big as 7500 and also scattered on a skeletal slope. At one point there was a gravel patch with Gibbaeum austricola which is commonly found on such gravel patches associated with ferricrete inselbergs.









4. MBB7504 Die Kop North (figs 12a to k). The plants are smaller and there is a more evident floribunda influence. I did not think that these plants were significantly different from the many populations discussed in Chapter 15 between Tarentaal and Stormsvlei.











Conclusions:
It should be very evident from all the information provided so far that more information is not producing a better picture of discrete elements. To the contrary and this reinforces the conclusion already mooted, that movement to a new view of “species” in the subgenus Haworthia is necessary, but is it possible?
Acknowledgement:
Mr Keith Spencer, DeHoop Nature Reserve.
Messrs Christine & Malcolm Wallace, Ziekenhuis.
Mr John Douglas-Hamilton, Ballyfar.
Mr Johan Groenewald, Klipfontein, Potberg.
Messrs Saartjie and Neil Neetling, Juliesfontein.
Messrs Adele & Vlooi DuToit, Die Kop & Frederickskraal.
Messrs Pet & Johannes Uys, Die Kop.
Messrs Sofia & Jurie Vermeulen, Tarentaal