
Introduction
I wonder. I have written so many words purporting to be my last that my credibility here too must be under stress. Two very recent articles of mine in Alsterworthia deal essentially with that issue, although they also cover the discovery of Haworthia mutica (Buffeljags) (= H. groenewaldii Breuer). They do not cover my subsequent thoughts on actually reading the description of this new “species” by Breuer, Marx and Groenewald. I hope that the present manuscript will explain why I reject this as a Latin binomial although anyone who is in the least familiar with my writing should already know. Spurred on by that discovery, I instigated a search in another area of the Buffeljags valley adjoining the Bontebok Park accompanied by Jannie Groenewald who informed me of what he had found in still another area I had long wanted to explore. So I instigated another search there too and again with Jannie. A discussion of these new finds is submitted to Cactus and Succulent Journal where I trust it will be published. The essence is already in Alsterworthia and this article is written to widen the readership, submit more pictures and maintain continuity with the 6 volumes of Haworthia Update that Harry Mays has been so conscientiously and determinedly publishing. This is all writing that may not otherwise have seen the light of day. I am personally extremely grateful for that as I have had a mania since writing my revision Haworthia Revisited and Update Vol. 1 (both Umdaus), to set the record straight and explore all the unknowns, or at least some of them.
The second area opened to me by meeting Jannie, was a farm southeast of Tradouw Pass, Heuningklip. In the Tradouw Pass, H. retusa ’turgida’ is found on the steep cliffs at the southern end. The road has been rebuilt since I first saw the plants and the population is greatly reduced from what it was. I simply do not have a picture handy to show what those plants are like, but that is not the need here and it does appear in Haworthia Revisited. Immediately south of the pass is another population of plants that puzzled me from the moment I saw them until now. In Haworthia Revisited there is a picture of one clone under J.D. Kobus Venter’s number 93/35. It is named as H. magnifica var. magnifica. The plants are present in a tension zone between Shale and River gravel, and vegetationally between what was Karoo Valley Bushveld and grassy Renosterveld. In April 1997 I collected seed and grew countless seedlings from there, which were all different. Only one picture, as MBB6666) is included in my long treatise on what I discussed under the title of “How to understand H. mutica var. nigra” inHaworthia Update 2.1:50. That picture is also under the name H. magnifica var. magnifica but it was not intended to stay that way. . It must be noted that my use of names in the Update volumes was not in a decision-making process so much as a learning and informative one. I read my conclusion to the treatise and note that I made no formal name changes. It is only as further field exploration yielded more and more information that I came to realize that H. magnifica and H. mirabilis, classification-wise, are inseparable. The Tradouw collection still worried me because of the close proximity of H. retusa ‘turgida’. These plants at Tradouw and all the things I referred to as H. mutica ‘nigra’ are in fact better understood as variants of H. retusa in the complexity of its relation to H. mirabilis.
This article thus covers exploration of an area between Swellendam and Heidelberg that I have neglected for too long. It is particularly applied to the problems of classification and naming that have persistently clouded my life and my relations with other writers. But as importantly to me is the unclouding of the way taxonomy and classification is useful and necessary for those interested at all in plants and how it is viewed by them.
Consideration and use of formal names.
Here I need to explain the naming style I now adopt. I have dropped the use of any rank below that of the species name. I do this because botany has no proper species definition and consequently species descriptions are just based on wild guesses about possible non-similarity and on the flimsiest of supposed character differences. The loosely used word “typical” is only truly useful in respect of the one plant dried as an almost unrecognizable herbarium specimen that is used to anchor the Latin name. The scientific ranks are aggregates and the terms variety and forma are so too. There is simply too much variation between populations and within populations to enable the certain identification of almost any plant that the binomial system suggests. There are no clear entities and ranks and this article should demonstrate that. There is a separate code of nomenclature that covers cultivars. Note that I do not drop names. I only drop the suggestion of status that additional names have, be they single clones or population references for all the different plants within them. I have used names in Haworthia for 70 years and intensively for the last 40 years and I am very familiar with the difficulties of any system. For the field and all the populations and plants I have seen, I can do no better than offer the list of names I updated in February 2009 in Haworthia Update 5.2:192. This, and the set of Updates, comprises a near total overview of the genus and effectively a revision. The attempted revisions by others fall short for me because they do not cover the range of populations that I have seen nor what history tells me, although they may include populations not seen by me. That is not strange because the area to explore is still vast. Another problem with even trying to be formal, is that which G.G.Smith encountered (Mrs L. Bolus did too) where several people were generating new names at the same time and trying to keep track formally with concurrent name changes would not have been, is not, possible. The point I should make is that in the last ten years, my field work has confirmed the predictive nature of my classification and these latest finds are in extraordinary agreement with that kind of expectation. What I have found essentially agrees with what I would expect. This is dramatically so for H. mirabilis based on a find by J. Jelimicky at Sandhoogte midway geographically and visually between H. mirabilis ‘bobii’ Hayashi (my nom. nuda ‘pilosa’ ,’velcro’, Steven Hammer’s ‘Shaggy dog’) along the Breede river, and a collection of mine further south in the DeHoop reserve, MBB7886 H. mirabilis, discussed (p11) and illustrated (p33) in Haworthia Update 5.2.
Prior to extending exploration in the Buffeljags area, I had also been down to the Haworthia populations in the lower Breede River area. This is where H. mirabilis seems to assume a wide range of forms and H. retusa ‘turgida’ and H. variegata’ are also present. I also, in the company of Kobus Venter and Lawrence Loucka, revisited Kransriviermond and Morning Star south of Heidelberg where H. floribunda is in the mix with H. retusa ‘turgida’ and H. mirabilis (in my broad new sense that includes H. maraisii, H. magnifica and H. heidelbergensis). Thus many of those images were very fresh in my mind when I went to Heuningklip southeast of Tradouw. Looking at all the plants and in editing my pictures I was truly impressed with the reality of continuity. The essence of H. retusa and H. mirabilis is in all those plants. A mobius strip is a strip of paper twisted and joined at the ends to produce a shape that only has one surface. What we have in the Southern Cape, and covered or touched on in this article, are interlinking mobius strips with one main surface as well as loose ends that present identifiable end-points. I tried once to illustrate the continuities between populations with single or multiple lines joining the different populations to express degrees of similarity. I can also visualize a pie-chart in which each population is represented by a circular chart with the contribution of main elements drawn as slices proportional to the strength of contribution. Table 1 is a schematic and very crudely suggested enumeration for such a set of charts. I have put the elements retusa and turgida as separate under an H. retusa heading for no other reason that I want to emphasize turgida as a variant at the two geographic extremes. One is eastwards where H. retusa ‘retusa’ is absent (Brandwacht, Mossel Bay) and one westward where it is present (in mutica, Hasiesdrift, Bredasdorp). H. retusa ‘turgida’ is present both east and west. But there are many populations contributing to the H. retusa pie in all the ranges of the two parts, from 0-50% thus making up the continuity from retusa to turgida.
Consideration and characters.
a. The pie chart.
Measuring the similarity of any two plants is extraordinarily difficult. Doing this for a population is even more so. Doing so for many populations, including those unseen in order to fulfill the predictive requirement of a proper hypothesis, is next to impossible. Hence all the disputation and name abuse. In table 1, I speculate an arrangement to illustrate the problem. The pie is constructed from the figures in the chart for just that set of genetic characters that are directly relevant to the set of plants in question. The pie is incredibly big and it should be understood that there must surely be pie-parts that spread their influence beyond the restrictions of the slices I conceive. So my table is restricted to the few species that are immediately relevant in respect of geographic association and direct interaction (by possible cross-pollination?). A pie-chart is drawn for each individual population. In the chart I assume that there is a set of genetic information that programs for H. retusa ‘retusa’. I have also suggested a different set for H. retusa ‘turgida’. If one considers only these two elements and all the known populations, my suggestion is that there is a set of charts that moves smoothly from one set to the next with no interruption. There is no discontinuity and the sets average 50:50 for the information. Thus it is one species and H. turgida is noted as H. retusa ‘turgida’.
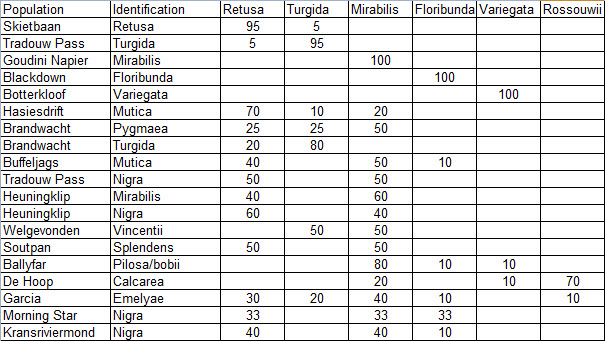
Table 2 is the complete set of pie-charts that should convey the possible relationships of the different populations and from which can deduce and understand to what degree H. retusa and H. mirabilis can be related. Also it will indicate the similarities between, and the difficulties of, trying to explain H. pygmaea in the east and H. mutica in the west. Both arise from the same genetic source. The problem is compounded by the infiltration of H. floribunda in the area midway between Heidelberg and Swellendam with the re-emergence of H. floribunda in the south at Potberg. H, variegata also enters the picture but from the south and also in the Potberg area. H. emelyae is the very probable combined entity entering the little Karoo. H. rossouwii is almost independent except in the very south-west at Bredasdorp and it may possibly link to other elements such as H. herbacea and H. reticulata.
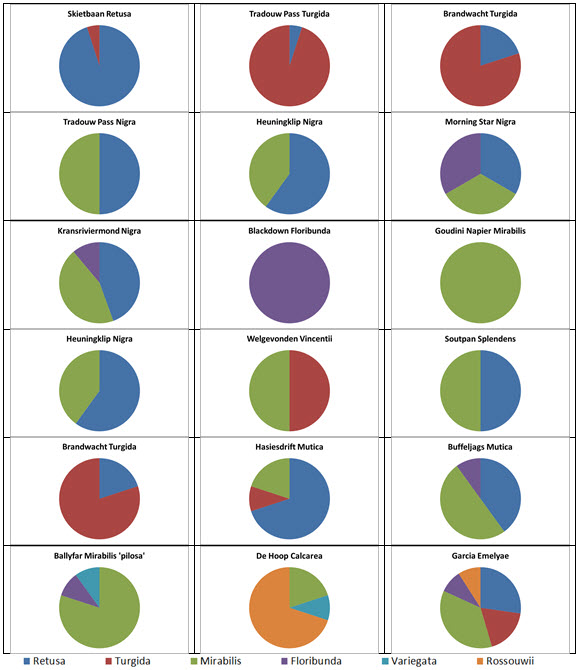
At Heidelberg there is interaction between an element that is a 50/50 retusa/turgida with a variant of H. mirabilis cf ‘heidelbergensis’. The latter name suggests that H. mirabilis is the general name for all the plants of that species, and ‘cf’ is an abbreviation for ‘compare with’ ‘heidelbergensis’ to refer to the small plants in populations previously regarded as independent species.
b. What really constitutes the pie.
The pie is the totality of the genetic material, the DNA that determines all life forms. It is the genes, the materials of characters and inheritance, and the repository of all genetic information. The pie is very large indeed and it is said that the DNA pie of an elephant only differs by 3% from that of a mouse; that of a man by 1% from a chimpanzee. In Haworthia one species from another ‘species’, I could guess at 0.001% or less regardless of whose concept I follow. This difference does also not reside in one consolidated slice that can be reduced to a single small pie as I have done. In DNA analysis, certain segments of DNA from cellular chloroplasts, mitochondria or the chromosomes themselves are isolated and the sequence of the paired amino-acid is analyzed. It requires a formidable statistical process to arrive at a two-dimensional tree (phyllogram) that illustrates a branching relationship with a measure of branch separation and lengths. This can perhaps also be used to generate pie-charts with an innumerable number of slices. I personally consider the phyllogram generated in the DNA sequencing process grossly inadequate.
c.) The characters.
This is extraordinarily difficult to specify no matter how obviously the plants seem to differ from one another. Number, size, shape, colour, arrangement, surfaces, spination and attitude of the leaves in the spiral rosette constitute the main things that determine how we see the plants. These are influenced by growing medium and growing conditions so these are complicating factors. Flowers are used in the same way as the leaves but far less so because they are so similar in all the species. Flowers are also not always present either which reduces their usage. Time of flowering is a major factor because it is linked to the reproductive process. It is also difficult to capture any firm picture of the flower because of the aging process in the 3-5 day life of the individual flower.
d.) The relevance here.
It simply is not necessary to parade the obviousness of this and the way in which will be attempted anyway in the discussion below. It is flowering time that may be critical beyond, or linked, to the facts of spatial geographic relationships of populations. It is traditionally flower structure that has formed the basis of classification of plants because of the reproductive significance and the role of reproduction in distribution of genetic material between plants and populations. Pollination of Haworthia seems to be primarily by a Solitary bee species. These bees make individual nests and are not communal like the Honey bee. They make tunnels in the soil as nests during the summer months. I have observed them as adults in the winter but then they are less numerous and less active. Few Haworthia flower at the height of winter and do so in two main time frames viz. spring and late summer. The implications are that breeding between isolated populations apart from any other consideration of compatibility, is a question of distance and time. A nest bound Honey bee can forage to 13km from its nest. A free flying Solitary bee can be speculated to cover a considerably greater distance. All these things play on the frequency and power of inheritance. Genetic drift is acknowledged as the genetic movement within a population away from the norm for a species because of isolation, while genetic flow is perhaps the holding together of similarity over geographic distance by a travelling pollinator. Thus the statistical probabilities of breeding across distance are impossible to derive.
I cannot quantify any of these things nor suggest any greater knowledge but this suggests to me, given the freedom with which the various species hybridize in cultivation and even in the field, we need to consider geographic relationships. In my extensive experience with other genera, it appears to me that a key consideration is if things grow together and maintain their difference, or not. Haworthia species are invariably in small isolated populations and seldom do different species grow intermixed. They are also influenced by geology of the substrate and particularly so because they favour the lowered competition from more vigorous and larger plants on the rocky sites or shallow soils they favour. That can be stated in the converse, Haworthia are favoured by conditions poor for other plants.
e.) Flowering time and growing together.
The existence as discrete species can be judged from the way in which populations share geographic space. I discussed this in some detail in Update 2.1:76. If two sets of plants grow close together and flower at the same time, while still maintaining obvious group differences, then they are most probably different species. If they flower at different times while still maintaining difference then the question of being different species is taken as certain. If the groups are separated and flower at different times while the differences are small and apparently within the experienced range of variation observed between populations, then it may be problematic. One would need to resort to the extended distribution and variability of the groups in question. Generally in Haworthia, species belonging in the same sub-genus do not share habitat and this is indicative of fairly rapid absorption of introduced genetic material (by pollinators). Hand-pollination and the ease with which plants of different species hybridize in cultivation suggest that there is practically no breeding isolation between them. In the following discussion, use is thus made of general distribution of species and proximity of populations in assessing to which species they best belong. It should be borne in mind that variability may be clinal. This means that the plants change along a geographic gradient. The direction of the gradient may also not be of increasing distance i.e. linear. It may also be circular so that ends of a gradient may meet again as two populations that appear wholly different, but linked by an extended set of other populations. A much more complex situation may actually exist where these clinal gradients may be multidirectional and thus involve more than one apparent species. My observation is that populations do differ and differences follow a pattern that becomes predictable i.e. gradual genetic difference over geographic area. This is not just a result of environmental gradient, but also linked to the question of genetic drift (the tendency of isolated populations) to move away from the general norm. It also results from the genetic flow between populations where the change of genetic material between populations at the distribution extremes is only through other populations.
f.) The species definition.
This is a subject I have covered many times. My view is that species are dynamic chaotic systems oscillating between different character states with time and with different rates of mutational change. Closely related species may not be recognizable on the basis of single character states and the spatial geographical distributions and local juxtapositions of groups of plants may be the only way to understand what the situation is and so make sensible decisions.
g.) Habitat geology.
I contend that species richness in South Africa is largely due to its varied geology, skeletal soils and comparatively mild climate. This present discussion deals with species that are associated with a geological phenomenon that is the Tertiary deposits of the Southern Cape. I am no geologist and am required to make the pretention I am one to explain why plants are distributed the way they are and what influences difference among them. In this case the vegetation at Buffeljags, Swellendam has been described (by Breuer et al. cited below) as on silcrete, and the general vegetation as by vegetation scientists as Ruens Silcrete Renosterveld. But there are two problems. The first is that vegetation classification is not infallible and I have never found it really useful in dealing with a wide range of species that occupy skeletal habitats. Secondly the substrate geology is definitely not simply silcrete nor is the clay soil either simply bentonite or kaolinite. There are many problems in the classification of clays. Bentonite in common terms seems to be the yellowish clay mined in large quantity from Heidelberg eastwards and there is a transition to ochre bearing clay. Bentonite may be aluminium, potassium, sodium or calcium dominated. As a montmorillomite clay it should have expansive cracking properties. Kaolinite is popularly regarded as the very white clay that is commonly seen under the rocky top layer of the sharp inselbergs that rise above the wheatfields west of Heidelberg. It seems to be less mineral dominated clay and does not have the expansive/contractive properties of bentonite. The distinction between ferricrete and silcrete seems to be that in ferricrete, iron oxides are infused into the petrified water deposited layers. Both the kaolinitic and bentonitic clays seem to be derived from the weathering of Bokkeveld shale that underlies the silcerte/ferricrete layer. It does not require any stretch of the imagination to see now why any one inselberg may be a little different from the next, and sometimes a lot different. What we do not know is how rapidly the landscape has moved from a broad white silcrete/ferricrete underwater plain to the remnant condition of that rocky plain today. Neither do we know what it has meant for the plant species that are linked to, and dependent on, the habitats generated in the erosion processes.
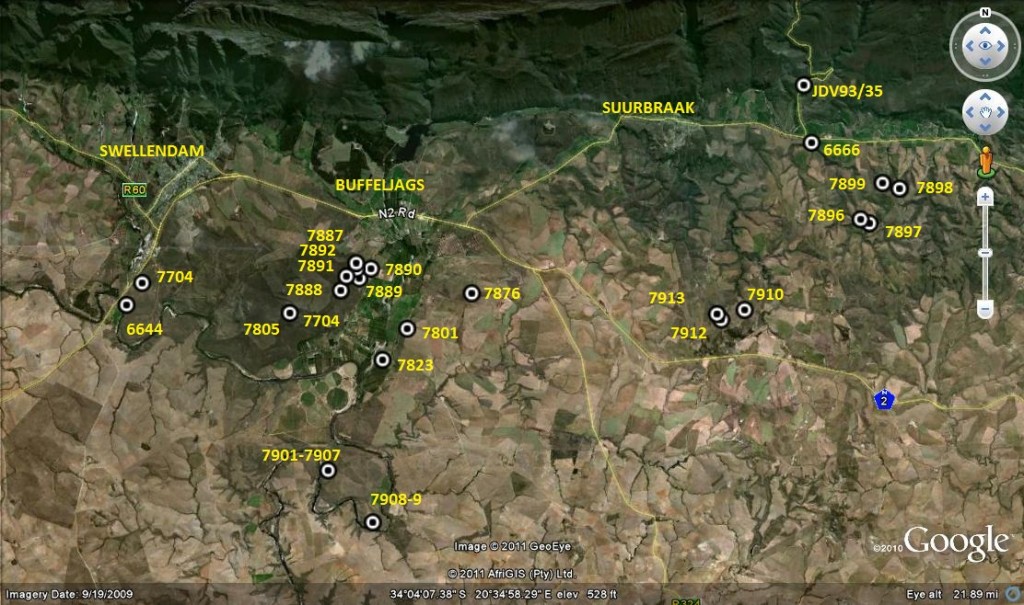
Map of Swellendam to Suurbraak area.
Map Legend – east of Swellendam.
1. MBB7899 H. retusa ‘nigra’. Heuningklip.
2. MBB6666 H. retusa ‘nigra’↔ H. mirabilis. S Tradouw Pass.
3. MBB7896 H. retusa ‘nigra’. Heuningklip.
4. MBB7897 H. retusa ‘nigra’. Heuningklip
5. MBB7898 H. retusa ‘nigra’. Heuningklip
6. MBB7888 H. mutica. Rotterdam.
7. MBB7889 H. mutica. Rotterdam.
8. MBB7890 H. mutica. Rotterdam.
9. MBB7801 H. mutica ‘groenewaldii’. Buffeljags.
10. MBB7741 H. mutica. Dankbaar.
11. MBB7892 H. marginata. Rotterdam.
12. MBB7891 H. minima. Rotterdam.
13. MBB6644 H. mirabilis. SW Swellendam.
14. MBB7704 H. mirabilis. Bontebok Park.
15. MBB7805 H. mirabilis. Bontebok Park.
16. MBB7823 H. mirabilis. Klipbult.
17. MBB7887 H. mirabilis. Rotterdam.
18. MBB7876 H. mirabilis. Disselfontein.
19. MBB7901 H. mirabilis. Crodini E.
20. MBB7902 H. mirabilis. Crodini E.
21. MBB7903 H. mirabilis. Crodini E.
22. MBB7904 H. mirabilis. Crodini E.
23. MBB7905 H. mirabilis. Crodini E.
24. MBB7906 H. mirabilis. Crodini E.
25. MBB7907 H. mirabilis. Crodini E.
26. MBB7908 H. mirabilis. Sandkraal W.
27. MBB7909 H. mirabilis. Sandkraal W.
28. MBB7910 H. floribunda. Rietkuil.
29. MBB7912 H. mirabilis. Rietkuil.
30. MBB7913 H. mirabilis. Rietkuil.
31. JDV93/35 H. retusa ‘turgda’. Tradouw Pass.
The populations.
Set 1 – THE TRADOUW PASS AREA (H. retusa ‘nigra’).
This first section deals with populations that complement those in the long discussion under the title “How to understand H. mutica var. nigra” in Haworthia Update 2.1:50. There I ended with the words…”.The habitat photo (of H. mutica Hasiesdrift) raises such strong images of H. retusa that doubt about the affinity of the var. nigra cannot easily be laid to rest.” Added were these words…”There the saga for the moment must rest.” What I had in mind was exploration that no one seems very anxious to do. This has now been done and I would lay the matter to rest as follows:-
1. MBB7899. H. mirabilis, Heuningklip. See figs 1-30 Fig. 1 is a scenic view of the area to show the countryside and the type of inselberg the plants are often found on. Fig. 2 is a closer picture of the erodible clay held in place by grass clumps at the base of which the plants often are. Occasionally they are simply out in the open sun. I term these places pressure bursts because water build-up in the soil causes collapse of the small banks and erosion can be quite fast. Several times I have found plants lying in collapsed soil. Such a soil bank near Heidelberg has retreated by nearly 2 meters in about 15 years albeit accelerated by road maintenance, but it does relate to any attempt to explain how much suitable habitat may have disappeared with time.

Fig. 1 A view looking north from Heuningklip to Tradouw. MBB6666 at arrow and MBB7899 at 6pt star – b 
Fig. 2. A photograph of the habitat of MBB7899.
Figs 3 to 30 is a selection of plants at the site where one is almost compelled to examine each plant separately and easily generate another 28 practically different images. Fig. 3 is of a plant with the surfaces roughened by small tubercles. SEM (scanning electron microscope) images of similar leaf surface often show incipient tiny spines on the tubercles and they appear as spines or hairs in populations like H. mirabilis ‘pilosa’ at Ballyfar, or H. mirabilis at Windsor Riversdale. This surface roughness and all its degrees is fairly characteristic of H. mirabilis but is does occur in H. retusa too. Plants illustrated in Figs 4 and 16 has one muttering Kruisrivier and Komserante, northeast and east of Riversdale respectively, and which I would have allotted to H. magnifica in the early period of my exploration no doubt have Breuer and Hayashi names. Now I simply say H. mirabilis and add the locality to place the plant to at least give it a geographic reality that is better than using the formal Breuer/Hayashi in hanging commas as I also suggest. Fig. 6 is apparently a seedling and demonstrates the leaf-shape of H. floribunda that led me to suggest that H. floribunda is a perpetuated juvenile form. Of course the evidence suggests to me that H. floribunda coalesces with H. mirabilis in this wider area although it does appear at Swellendam to the west of my map as the large form and at the extreme SE corner of the same map as a convincingly normal form. Fig. 9 demonstrates longer pointed leaves and Fig. 12 short more obtuse leaves. Fig. 17 is of a plant that has relatively smooth leaf surfaces rather like H. retusa and the leaves are conspicuously lined. In Fig. 20 there is a suggestion of muticate (without a point) leaves. Fig. 22 is again quite strongly lined. Fig. 22 shows a leaf with a single line. Fig. 25 demonstrates what is termed rotate leaves where the points of the leaves point around as though to encircle the plant. Fig. 28 made me recall plants of H. mirabilis at Melkhoutrivier in the south west near Malgas and further to Ballyfar. The flowering time is late summer.




























These images can now be compared with MBB6666 at Tradouw Pass to establish if my assessment is reasonable. My contention is that this MBB7899 population is midway between the other three Heuningklip populations I am about to illustrate and Tradouw Pass. This similarity is repeated widely and eventually contributes to my contention that H. mutica has been generated by drift away from its two contributing parent species viz. H. mirabilis and H. retusa.
2. MBB6666 H. mirabilis. Tradouw Pass. See figs 31-71. This population is at the mouth of the Tradouw Pass and relatively low-lying in respect of the previous population. The habitat is quite different being partially coarse river gravels, weathered shale and at one point is an un-weathered shale ridge where the plants are wedged in the cracks. The habitat is of course very different to the steep sandstone cliffs within the pass itself where H. retusa ‘turgida’ occurs. I initially thought these plants fitted with H. magnifica. A decision derived from a very small sample available at the time. Now I looked for the images and photographs in my records under H. mirabilis where they should have been since my inclusion there of ‘magnifica’, and alsounder H. retusa where I found them in the folder ‘nigra’. The flowering time is late summer. In this respect similar to the previous set MBB7899 H. mirabilis but the plants are more robust. Figs 31-45 were taken in 2009; figs 46-53 in 2011 and figs 54-71 are from seedlings grown by myself. I tried to reduce the pictures and eliminated a few. But I really think they all need to be presented to show again how serious the difficulty of circumscribing them as a single recognizable key-able entity really is. While looking at my pictures I seriously thought that the set taken in 2011 differed from those taken in 2009. The cultivated plants of course are not directly connected to their source and the variation is self-evidently wide. But it is equally so in field plants. There are the smoother surfaces of H. retusa and some very rough surfaces for H. mirabilis. My decision became that they fitted best with the ‘nigra’ populations where my organizing had taken them despite flowering time, but here MBB7899 shows absolute geographic position between H. mirabilis and MBB6666. I consider this key evidence for the basic continuity for H. retusa and H. mirabilis that I have conjectured elsewhere and I would not be able to defend an argument that MBB7889 plants belong better in H. mirabilis and this means MBB6666 belongs there too. I would simply dispute that there is no better solution so stick with history as far as possible and that means H. mirabilis. If Breuer or Hayashi have a name for a variant from this population intended as an aggregate name, I recommend the use in inverted commas. I would actually like the use of the equilibrium diacritic of a double arrow pointing between two names viz. H. mirabilis↔H. retusa ‘nigra’, although this available symbol is not quite the thing.

Fig. 31 6666 H. retusa ‘nigra’ 







































I come to this vexing question of “typical” that is bandied about even by myself when I liken population to population. There is the same range of variables in MBB6666 noted for MBB7899, but the plants do not darken as much and they are larger. So we need now to look at the other three Heuningklip populations asking what significance is attached to their presence in exactly the same habitat as MBB7899, but geographically separated and by perhaps 500m and 2km respectively. The plants are so variable that it is not possible to arrive at a characterization that separates any two plants, or any two populations in any geographic set for that matter.
3. MBB7896 H. retusa ‘nigra’, Heuningklip 1. See figs 72-89. The plants in this and the next two populations were flowering in September as does H. retusa and they are relatively large, green in colour and generally smooth. Of course both vary enormously due to both intrinsic genetic difference and extrinsic local situation. One plant within an older decaying grass tuft was nearly 200mm diameter, and a healthy green colour, another plant in the open white clay was less than 50mm diam. and dark in colouration.

Fig. 72 7896 H. retusa ‘nigra’ 















4. MBB7897 H. retusa ‘nigra’, Heuningklip 2. See figs 90-105. The first two pictures show the general landscape and the local habitat where the plants occur. The clay is highly erodible and often there are no plants at all in the clay. Some of the plants have very acuminate pointed leaves while in others they may be relatively obtuse and rounded.

Fig. 90 The countryside looking towards the Tradouw Pass. 
Fig. 91 The ‘white clay pressure burst’
5. MBB7898 H. retusa ‘nigra’, Heuningklip 3. See figs 106 -132. The great variability is again evident and in this population the plants were observed to be generally without much venation or lines as in fig.112. Then one plant, fig. 119, was so striped as to stand out from all the populations seen.

Fig. 106 Silcrete hilltops

Fig. 107 7898 H. retusa ‘nigra’. 
























Set 2 – THE BUFFELJAGS AREA (H. mutica).
This second section follows a recent manuscript submitted to Alsterworthia yet to be published entitled “Some last closing thoughts” written in the hope of finding the closure I seek and that will not come. That article is about a population that has been named as a species viz H. groenewaldii, in an article authored by Breuer, Marx and Groenewald (Alsterworthia 11.2:15) that I only saw after the submission of my manuscript. I also only saw the article after the exploration I would have asked for to verify opinions. The description seems to have been prompted by Marx who refers to similarities to H. springbokvlakensis and to H. magnifia var. atrofusca and raises many issues that my manuscript does not cover. I simply find the whole discussion unrealistic and especially so when no further exploration was done to examine anything. One should refer to Breuer’s “World of Haworthias Vol 1” to see that the naive promises in the foreword and introduction of exploration and field study, was never achieved. It is true that the plants have an unusual surface texture and it is also true that the habitat is quite unlike any other H. mutica occupies. But it is Marx’s comment that disturbs me viz. “Unfortunately Bruce Bayer keeps on refusing to see it as something new and calls it simply H. mutica ‘Buffeljags”. This is a distortion and misrepresentation, and whose misfortune this is, not specified. My reticence is reflected in the preceding discussion and Gerhard does not seem to follow, or agree with, what I have been writing about and trying to explain in this general context for so many years. In fact in an unpublished (to my knowledge) manuscript he dismisses all my writing as “pretentious” that no doubt it is, because I have never fancied my self as a taxonomist in the tradition of taxonomists. I was as fascinated as Marx by the plants and am even more anxious than he is to see just how they connect to the probable geographic counterparts in the context of the species definition that I apply. I was also very curious to know what the connection to H. mutica and the H. retusa ‘nigra’ issue, only now laid to rest in the preceding discussion, actually might be here. This is why I use the word “instigated” in reference to exploration in the very valley where H. mutica ‘groenewaldii’ occurs and to the obvious habitat for similar plants. It is surely unfair iniquitous to so demand that I see this population as “something new” in an article that rather disregards the history of the subject and what I have been trying to understand of it.
I also recently submitted a manuscript to Cactus and Succulent Journal (US) where I wrote of “H. groenewaldii Breuer”… “I consider it to be generated from the interaction of H. mirabilis, H. mutica and H. floribunda. I attach no special importance to the fact that it flowers, contrary to H. mutica, in February/March. This is because I have observed many hybrids between patently different species despite a seasonal difference in flowering time”. I have in many places discussed the problem of “hybridization” and it must surely be well-known from my writing that H. floribunda merges into H. mirabilis. Less well-known will be the fact that evidence for H. variegata doing the same is also available. In respect of floral characters, I wrote a chapter in Haworthia Update Vol.3.2:88 explaining the problems of using floral morphology for species recognition. I would challenge Breuer et al. to produce comparative results from studies of the flowers in any of these problematic species to demonstrate difference. Regarding the curious shiny leaf surface, I refer to Haworthia Update Vol.2.2:141, Chapter 15 where I discusselectron scanning photographs of leaf surfaces. This also demonstrates how problematic leaf surface is. It would be tiresome indeed to parade all the evidence in support of these contentions of mine. In Haworthia Update 5.2:108 I also briefly discussed H. mutica from Buffeljags (‘groenewaldii’) and submitted a large number of photographs explaining the habitat and what the possible connection was to the H. retusa ‘nigra’ conundrum. I may also have written about the issue in Haworthiad. Suffice to say that the authors of H. mutica ‘groenewaldii have certainly not checked or considered “all the facts” as the principal author claims. The very opposite seems to be the case.
Considering my oft repeated observation that geological substrate plays a major role in shaping species appearances, the Breuer, Marx and Groenewald description of the habitat for H. mutica ‘groenewaldii’ is a little misleading (see considerations above). Nowhere else is H. mutica on identical substrate and this would just support my contention of ecotypic differentiation that does not justify a Latin binomial. There is considerable doubt about the actual difference between these river boulder situations and the widely distributed terrace gravels elsewhere in the area, notably the Bontebok Park near where the following populations were found:
6. MBB7888 H. mutica, Rotterdam 1. See figs 133-169.

Fig. 133 7888 H. mutica. 



































7. MBB7889 H. mutica, Rotterdam 2. See figs 170-188.

Fig. 170 7889 H. mutica. 













Fig. 185 



8. MBB7890 H. mutica, Rotterdam 3. See figs 189-191
The three populations above are separated spatially by several hundred meters and they are all on remnant river gravels on a more gently sloping bank than the Buffeljags east bank. The vegetation is closer to conventional fynbos as in the adjoining Bontebok Park, but grassy. The pictures are self-explanatory and the leaf surfaces of the plants are also generally as for the Buffeljags population. What is significant is that I cannot say I saw a single plant in these populations that resembles the one chosen and pictured as typical for this highly dubious species, H. groenewaldii. It makes more sense to me to identify plants from these populations by collecting number than to generate a supposedly descriptive Latin epithet from an imagined typical representative.

Fig. 189 7890 H. mutica. 

9. MBB7801 H. mutica ‘groenewaldii’, Mullershof, Buffeljags. See figs 192-215
The following set of photographs has surely seen the light of day somewhere and I repeat them because as far as I am concerned they are H. mutica with the shiny surface claimed for “groenewaldii’. Dankbaar is some 16km west of these Buffeljags populations and the substrate is Table Mountain Sandstone. The flowering time is just normal for H. mutica. Fig. 215 is a photograph of a plant apparently collected by Jannie Groenewald and it is quite uncharacteristic of the population. I would suggest that it is a product of hybridization with H. mirabilis in the vicinity (eg; Klipbult).

Fig. 192 7801 H. mutica ‘groenewaldii’. 






















Fig. 215 7801 H. mutica ‘groenewaldii’.
10. MBB7741 H. mutica, Dankbaar. See figs 216-235.
My observations in respect of these two sets of plants confirms my widely written convictions about Haworthia taxonomy and the state of classification in the absence of a definition for the concept of “species”, and the intrinsic value of Latin binomials for rational and sensible communication. Of course this Dankbaar population is not identical to other H. mutica populations and I am not driven by fixation on any one character that defines this species. It is a geographic set and the various sets are not simply separable. But I am not finished yet.

Fig. 216 7741 H. mutica. 

















Set 3 – THE BUFFELJAGS AREA (Robustipedunculares).
The western Buffeljags area also produced a population of H. marginata and one of a very curious H. minima, that both complement occurrences in the Bontebok Park a little further west. The conventional small, blue-green H. minima is present in the park as well as an unusually green population.
11. MBB7892 H. marginata, Rotterdam. See figs 236-243
This population was quite exceptional and one of the strongest populations known to me. But the plants were in flower in September whereas the flowering time for the species is normally late summer; and the flowers and seed capsules exceptionally large. I do not consider that these factors command another Latin binomial.








12. MBB7891 H. minima, Rotterdam. See figs 244-252 and Bontebok Park figs 253-4. These were also extraordinary plants, larger than those seen in the Bontebok Park population where the plants were also green viz. figs 253-4. Fig. 252 confuses the issue because, while the other plants seemed alike to H. marginata and distant hybrids, this one plant appears to be a recent hybrid. This occurrence cannot be overlooked in respect of other introgressive (or seemingly introgressive) populations of Robustipedunculares species. The plants were not in flower and again we have a suggestion or direct evidence of hybridization across a difference of seasonal flowering.

Fig. 244 7891 H. minima. 








Fig. 254 sn H. minima, Bontebok Park
Set 4 – THE GREATER BUFFELJAGS AREA (H. mirabilis).
This is a set of 15 populations of H. mirabilis that occur east, west, south north of the H. mutica ‘groenewaldi’ related populations. I exclude from the discussion and illustrations the several populations of H. floribunda ‘major’ populations to the northwest, while I repeat the statement that H. floribunda contributes to the general gene pool of H. mirabilis and together with H. retusa, they all contribute to the gene pool of H. mutica ‘groenewaldii’. The huge presence of populations more readily assignable to H. mirabilis (sensu lato – in the broadest sense) is used by me to validate my opinion that the “mutica” elements draw most genetic resource from the H. retusa element, and hence why I refer those plants to H. mutica.
The sequence of populations 7901-7908 is located in a horseshoe of the Breede River, southwest of Buffeljags in very promising habitat. Only H. mirabilis was seen. I perhaps need to point out that these populations would have been extraordinarily difficult to name under any previous dispensation of mine where I was still trying to uphold H. magnifica, H. maraisii and H. heidelbergensis as species. These I have since merged in H. mirabilis and I reject the introduction of a group name like “aggregate” as Breuer seems to have done, as a complete fabrication that does nothing to solve the problem other than to highlight it.
The following 6 populations are all associated with tertiary and river gravels. There are differences between most of the populations but of an order that I cannot articulate. It is also not possible to really encapsulate the appearances of the plants in the two photographs for each population that I submit. At the same time it does not seem to me possible to make any communicable distinctions.
13. MBB6644 = 6860 H. mirabilis, SSW Swellendam. See figs 255-256.


14. MBB7704 H. mirabilis, Bontebok Park. See figs 257-258.


15. MBB7805 H. mirabilis, Bontebok Park.See figs 259-260.


16. MBB7823 H. mirabilis, Klipbult. See figs 261-262.


On selecting these two pictures I was reminded of my observation that there was a significant H. floribunda appearance to the plants. I had believed that I had seen H. floribunda in the Bontebok Park nearby many years ago but subsequent exploration has only produced H. mirabilis albeit with some similarities to H. floribunda. My reaction was to suspect that H. mutica ‘groenwaldii’ most probably also then needed to be considered as a product influenced by H. floribunda.
17. MBB7887 H. mirabilis, Rotterdam. See figs 263-264.


18. MBB7876 H. mirabilis, Disselfontein. See figs 265-266. These plants were a little more robust than the others in this set and rather similar to MBB7901.


The following populations are almost entirely in Bokkeveld shale but on a very close interface with tertiary gravels. In some places there was also some wind-blown sand.
19. MBB7901 H. mirabilis, Crodini 344, S Swellendam. See figs 267-278.












20. MBB7908 H. mirabilis, Sandkraal, S Swellendam. See figs 279-282.




Set 5 – BACK TO TRADOUW PASS – AS IT HAPPENED.
In the process of compiling this manuscript I was bothered by the ever present question of what is still out there that might influence the outcome. My wife, Daphne, was planning an outing to see chameleons at Swellendam while simultaneously contacting landowners for me around the Potberg. Always anxious to get two birds with one stone, I asked her to find out who owned the land west of Heuningklip that we had just explored with Jannie Groenewald. The result was that we ended up spending a day with Odette Curtis of Custodians of Rare and Endangered Wildlife, south of Suurbraak and finding three populations that, for me, put the proverbial cherry on the pie.
All three were on the slopes of silcrete inselbergs and almost the last of this habitat westward from Heidelberg to Buffeljags. The first inselberg was different to the second and from the Heuningklip habitats because it was dominated by Aloe arborescens and A. ferox. Aloe brevifolia was also present in an unusually small form. The habitat was very impacted by grazing animals and there is an outside possibility that H. retusa ‘nigra’ may once have been present although I personally doubt it. To our surprise we found…
21 MBB7910 H. floribunda, Rietkuil, Suurbraak. See figs 283-288.






There were many plants but severely trampled and disturbed. There survival was obviously aided by the fact that much of the plant is below soil-level. But my experience of this species is that it struggles to maintain its identity in the presence of either H. mirabilis or H. retusa. They never share immediate habitat. There was no hybrid present and this also adds to the problem of interpreting flowering time and distance apart from other species as part of the process. The plants were not the large green variant of the species present to the west at Appelbos, Swellendam; and the leaves were both with and without marginal spines. Eastwards I know the species to be close only at Blackdown north of Heidelberg and at Goedverwagting south of the N highway about 15km east southeast. It definitely seems to be absorbed into H. mirabilis west and north respectively of those two populations. So while this is a slight contradictory element of surprise, it is the next two that really excited me. These were again inselberg habitat but without the strong silcrete upper layer of the former habitat and without the Aloe element. The second ‘population’ was about 200m from the first and comprised four groups of plants along a longer north-facing ridge.
22. MBB7912 H. mirabilis, Rietkuil, Suurbraak. See figs 289-306


















23. MBB7913 H. mirabilis, Rietkuil, Suurbraak. See figs 307-331
I am very hard-pressed to say much more than that these plants met all my expectation for the very variable set of plants and populations that I ascribe to the one single species H. mirabilis. Who could possibly articulate the differences of the many populations of this species from each other and from this one? Variable becomes a done-to-death word. The individual plants, in two populations quite unlike any others I have seen, were generally robust with large specimens up to 100mm. diam. The colour is dark, although green when shaded within or under grass. The leaves were usually sharply pointed but there was always evidence of one or two leaves or odd individuals in which the leaves were muticate i.e. without points. In some plants the leaves were notably lined in others not. The colour also varied from chestnut to deep reddish-brown when the plants were more exposed. The surface texture was generally tubercled but there was evidence of smoothness of the leaves in many plants. Flowering time was obviously late summer gauged by the absence of flower at the time of the visit viz. September. Except for one oddity. This was a single plant with a weak flower spike. I photographed this for the useful information the flower-bud provides. Where Breuer et al. claim to have considered all the facts, this is a character that is conveniently overlooked when claims are made for the significance of floral characters when paraded to substantiate yet another claim to species significant difference. The bud-tip in all the Southern Cape species of subgenus Haworthia with the exception of perhaps H. rossouwii have this very obvious “fish-tail” bud tip. The tips of the upper two petals are flattened and spreading to resemble a fish’s tail. It is most marked in H. herbacea and H. reticulata where the bud tip curves downward from the semi-erect position of the body of the flower and then up again. I am fearful that my inability and reluctance to consider these plants as “new” may be the motive for someone to rush forward and apply a Latin binomial.
My opinion is that these two populations mirror the paradox in the east where H. pygmaea ‘fusca’ is nearer to H. retusa ‘retusa’ andwhere H. retusa ‘turgida’ is nearby. H. mirabilis ‘splendens’, in the doubt about its affinities, in my opinion remains in the context of H. mirabilis. This is a conclusion I reached after exploration of the area between Riversdale and Albertinia where the same elements noted in this article repeat themselves. I do not for one moment concede that better explanation is available by resort to a multitude of Latin binomials and by an artifact like “aggregate”.
























Conclusion.
I have been deeply disappointed that so little progress on the classification front has been made among the fraternity writing about Haworthia. In communication with Lawrence Loucka, he wrote…”You might consider saying that a genus revision is needed and that until amateurs depart no proper botanist will make the investment. Until then chaos will rule, names will proliferate.” This is a useful and insightful comment and generally summarizes what I have often said. Unfortunately the position has been rather exacerbated when proper botanists have made their presence known and I can recount many instances to support this statement. My own view has been that botanists have not been able to confront genera that are so publicly and popularly known, and garlanded with names. Thus amateurs, including myself despite at least an iota of formal training in science, simply have a field day generating names like rabbits out of a hat.
Unless some proper thought is given to this whole question of what the names are supposed to mean apart from catalogue items, taxonomy (classification) will never lift out of the disinterest, disrepute and scorn that collectors justifiably seem to have for it. Latin binomials do have a real value and to demean this by one-dimensional derivation and rationalization is intellectually fraudulent. My writing is replete with examples of the variability and interaction of populations over large area. Taking a single population and one flower, and often a single plant, to generate a species description and a Latin binomial is grossly simplistic.
The one change that this article generates is the confirmation that the population MBB6666 from immediately south of Tradouw Pass is better identified by name as an H. mirabilis variant, and not as H. retusa ‘nigra’. MBB7801 at Buffeljags and now also at Rotterdam, remains as H. mutica and the epithet generated by enthusiasts can be added to make the trinomial H. mutica ‘groenewaldii’. For formality, the name for the form illustrated as typical would be H. mutica ‘Groenewaldii’. A few additional names will be needed for similar clones and a lot more for the cultivars that will be derived from this population set.
My conclusions here make no substantial departure from anything that I have observed or written over 50 years. While the subject has been comprehensively covered in the field and in writing, there is a huge amount of exploration that still needs to be done if the genus is to be still better known. My prediction is that the answer will conform very closely to the species-list I finalized in 2009 (see Alsterworthia 5.1:192). The disclaimer is that botanical classification may eventually reach the point where the real nature of species and their purpose comes to be known. In that case what we now may see and persuade ourselves are discrete species, are not. Instead these highly complex chaotic systems may in fact be quantum units of consciousness and whole elements such as some genera and sub-genera we now recognize will be the species.
Acknowledgement.
I want particularly to acknowledge Jannie Groenewald whose interest and enthusiasm are boundless. Then Allan Jeptha, an environmental law practitioner with great vision for the Suurbraak community; Odette Curtiss, a consultant for the conservation of Renosterveld vegetation; Lawrence Loucka whose patient guidance and comment I value and who generated the set of pie charts; Kobus Venter and Steven Hammer who have always been supportive and contributory; Max Coetzee of Felix Unite and Samie Lategan of Buffalo Breeding Programme for access to Sandkraal West and Crodini East; Jaap Viljoen for his contribution; my wife Daphne, who trusts me and whom I deeply trust.
Dear Sir, Great article for interrelationship justification.
regards
Soumen Aditya