
M B Bayer, PO Box 960, Kuilsriver 7579, RSA.
The objective was to explore some likely habitats previously observed at Van Reenens Crest and nearby. We extended the scope to include further exploration for Haworthia mutica as I am still questioning the place of this species in the greater scheme of things. Thus here are four sets of populations that I report on viz. H. mirabilis, H. retusa ‘nigra’, H. floribunda and H. mutica. See maps Figs 1 and 2 for geographical position.

Van Reenens Crest 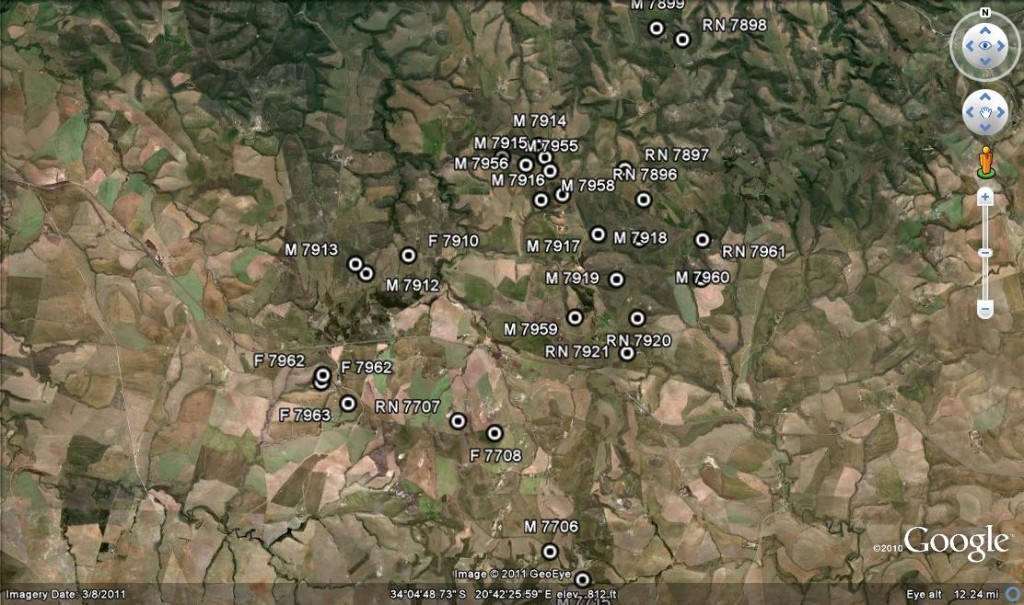
Fig. 1 Van Reenens Crest. M = H. mirabilis, RN = H. retusa ‘nigra’, F. = H . floribunda. 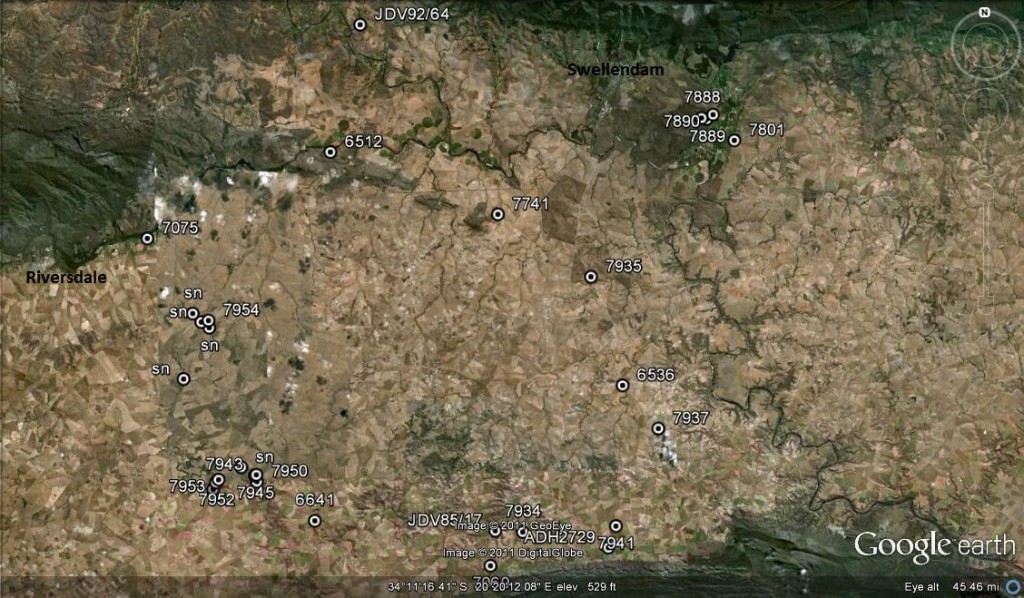
Fig. 2 Known distribution of H. mutica
H. mirabilis.
Set 1. MBB7955 Van Reenens Crest – mid west. (figs 3-11)
The plants are moderate in size to about 50mm diam. and growing on an erosional slope below tertiary gravels where there is white clay. They vary from densely tubercled to relatively smooth.

Fig. 3 7955 Van Reenens Crest 






Set 2. MBB7956 Van Reenens Crest – west 7955. (figs 12-34)
Despite apparently favorable habitat these plants were about 300m west of the previous set. Variation is again apparent in respect of presence or absence of surface roughness. Generally there are plants in which some leaves are without a point (see Fig 1) while others are acute tipped (mucronate – with a short hard point) or even have a long end-awn. Margins may be smooth or very lightly spined. This seems to be more apparent in young plants.

Fig. 12 View looking east to Set 1.

Fig. 13 7956 Van Reenens Crest 




















Set 3. MBB7957 Van Reenens Crest – west 7956. (figs 35-41)
Again the plants are very variable with dense surface tubercles to virtually none. The leaf shapes vary and in fig.39 that appears to be a group of seedlings there is a remarkable series of different leaf ends and margins. In some plants there is distinct spination of the leaf margin.

Fig. 35 View looking south to Tradouw Pass

Fig. 36 7957 Van Reenens Crest 




Set 4. MBB7958 Van Reenens Crest – south west 7957. (figs 42-51)
Fig. 43 shows two rosettes of a plant with a “crocodile-skin” surface. The surface tubercles seem to be flattened and the leaves are mucronate. The following photograph, fig.44, is of a plant with acuminate leaves ending in long end-awns, and the surfaces are relatively smooth – retusoid? Unlike the previous records for the area that are north-facing, this is a south-facing “pressure-burst” of white clay. A nearby habitat of vertical shale that might have been considered favorable for H. mirabilis elsewhere was devoid of plants.

Fig. 42 7958 Van Reenens Crest 








Set 5. MBB7959 Van Reenens Crest – west homestead 7956. (figs 52-64)
Figs 53-55 are smallish plants with rounded leaf ends and relatively opaque surfaces. Fig.58 is remarkably like H. retusa, say, at Pienaarsrivier with very awned leaf ends and a high degree of translucence in the leaves.

Fig. 52 MBB7958 View to MBB7957 a. Daphne inspecting, Lawrence checking the GPS.

Fig. 53 7959 Van Reenens Crest 










Set 6. MBB7960 Kruiskloof, E Van Reenens Crest. (figs 65–78)
This population departs from a very broad norm for Van Reenens Crest and picks up the continuity with populations north, east and south. This is most noticeable in the surface texture where the tubercles are smaller and denser. The leaves can be highly rotate, see fig.68. A feature seen occasionally in the Van Reenens Crest populations and as far afield as the Diepkloof (Malgas) area much further south, is the occasional plant with a very stellate (five –pointed star) arrangement of the leaves that are quite horizontal see fig.70. Among the many oddities is the plant in fig.72 with floribundoid leaf ends and a leaf surface where the tubercles seem to be flattened. In fig.73 the tubercles seem to be spined. In fig.75 tubercles are almost absent.

Fig. 65 7960 Kruiskloof 












H. retusa ‘nigra’
Set 7. MBB7961 Kruiskloof, E Van Reenens Crest. (figs 79-90)
These plants reinforce my perception that H. retusa and H. mirabilis are uncomfortably closely related. Figs 79 and 80 have the same general form as H. mirabilis ‘sublineata’. Fig.82 appears to me as possibly a hybrid as leaf opacity is deeper and there are marginal spines.

Fig. 79 7961 Kruiskloof 


Fig. 82 







H. floribunda
Set 8. MBB7962 Niekerkshek. (figs 91-99)

Fig. 91 7962 Niekerkshek 







Set 9. MBB7963 Niekerkshek. (figs 100-111)
The plants vary in size and to some degree in colour. This variation in colour from a deep purple to dark grey is very evident in fig.107. The leaves are much flattened and an occasional leaf is pointed. The plant in fig.102 is unusual in having white spots on the leaves. Marginal spination can be absent or very coarse. The leaf surfaces show no venation or translucens, as is expected in this species.

Fig.100 View looking south to Potberg.

Fig. 101 7963 Niekerkshek 






H. mutica
Set 10. MBB7950 DeDraai Klipbankskloof. (figs 112-123)

Fig. 112 – 7950 Die Draai, E Klipbankskloof 










Set 11. MBB7951 Klipbankskloof West 1. (figs 124-126)

Fig. 124 – 7951 Klipbankskloof West 1 

Set 12. MBB7952 Klipbankskloof West 2. (figs 127-129)

Fig. 127 – 7952 N Klipbankskloof West 2 

Set 13. MBB7953 Klipbankskloof West 3. (figs 130-144)

Fig. 130 – 7953 Klipbankskloof West 3 View westwards 
Fig. 131 – 7953 N Klipbankskloof westward view of habitat

Fig. 132- 7953 N Klipbankskloof West 3 











Set 14. MBB7954 Rondeheuwel. (figs 145-174)

Fig. 145 View looking northwest to Riviersonderend

Fig. 146 7954 Rondeheuwel 
Fig. 147 

























Fig. 173 
Fig. 174
Set 15. JDV92/64 Sanddrift, Drew (figs 175-187)

Fig. 175 JDV92-64 Very localised.

Fig. 176 JDV92-64 Sanddrift 










Set 16. JDV85/17 Haarwegskloof (figs 188-213)

Fig. 188 JDV85-17 Haarwegskloof 























There is a very wide range of variants among these 7 population sets. Leaf colour and shape vary enormously and it can only be said that generally the leaves are muticate. Almost invariably any one plant has a leaf or leaves that are also mucronate and in some cases even awned. The leaves may be abbreviated and short and squat or drawn out and elongated. Venation is also varied from many to few or only a single vein on the upper leaf surface. These veins may be straight and clearly separated or they may anastomize and link togther in a reticulate pattern. I did include two pictures of flowers, figs 173 and 174 just to indicate that simply taking two flowers at random one can see obvious differences.
Attention can be paid to figs 145 and 146 of Rondeheuwel where I consider these two plants indicate hybridization with H. mirabilis that does occur about 1km to the northwest. Fig. 147 shows a plant with a rather wrinkled surface.
Conclusion
My opinion is that consideration has to be given to the odd way in which the “species” do not share habitat. It is extremely difficult to rationalize an ‘average’ plant for each population and so establish any norm for proper comparison. The populations vary by degree. It is also clear that if one examines the distribution of the species across the study area, it can be observed that H. mirabilis where it occurs with H. retusa ‘nigra’(at Heuningklip and Kruiskloof in the east), is not the same as plants to the west where H. retusa ‘nigra’ is absent. Consideration has to be given to absorption into H. mirabilis. Why H. floribunda should make a re-appearance at Rietkuil and Niekerkshek in the west, when everything points to its absorption into H. mirabilis in the east at Skeiding, is a mystery. The fact that at Niekerkshek it also has radically different forms to those found anywhere else adds to the difficulty.
I will conclude by acknowledging that the populations of H. mutica are all summer flowering but I do not think that this is as significant as the authors of “H. groenewaldii” will contend. The Van Reenens Crest complex demonstrated dramatically that H. retusa and H. mirabilis share a very close common history and an equally close contempory relationship. My conjecture is that H. mutica is linked to that relationship in the same way the complex of populations are that I assign to H. pygmaea. It is painfully obvious that if floral differences that are so easily detected within populations, expecting to find more evidence in the flowers than exists in the vegetative structures across the genera, and especially the subgenus Haworthia is an absolute pipedream. Floral differences between species are minimal and if anything will provide support for a reduction of species rather than add to the. If any small difference is suggested to separate species, then equal weight must be given to similarities that should then unite species. The contention then that flowers are offering characters that I as an author have ignored, while already wholly untrue, is additionally nonsensical.
Acknowledgement
Mr and Mrs Trevennan Barry of Van Reenens Crest. Mr N Swart of Poststal, Mr J. duToit of Tarentaal and Wydgelee, Mr W duToit of Klipbankskloof, Mr J. Mathee of Klipbankskloof West, Mr Vlooi du Toit of Die Kop, Mr M Mynhardt of Haarwegskloof, Mr and Mrs Nelius Smith of Volmoed, Rondeheuwel., Mr Adrian Steyn of Wankie Boerdery. Thank you to Kobus Venter and Lawrence Loucka for company and shared observation.
Pingback: Van Reenens Crest - Haworthia Updates