
Printed in British Cactus and Succulent Journal 5:13 (1987)
Part 6. HAWORTHIA MACULATA (V. Poelln.) Bayer.
M. B. Bayer, National Botanic Gardens of South Africa, Karoo Gardens, Worcester.
Haworthia maculata (V. Poelln.) Bayer. Haworthia Handbook: 130 (1976); New Haworthia Handbook: 43 (1982); J. W. Pilbeam, Haworthia and Astroloba: 89 (1983). H. schuldtiana var. maculata v. Poelin. in Feddes Repert. Spec. Nov. 49: 25 (1940). Type: Cape; in the neighbourhood of Worcester, Swellendam, Bredasdorp and Caledon. Major H. Venter No. 6a. No specimen preserved. Neotype: Cape- 3319 (Worcester): Worcester District, H. Venter No. 6a in G. G. Smith 3912 (NBG)
Rosette stemless, 30-70mm in diameter, up to 60 leaves. Stem thick, white-fleshed, non-fibrous, 1/3 diameter of rosette, never elongate, slowly proliferous from base. Roots thick, white-fleshed, non-fibrous. Leaves erect, spreading, slightly incurved at tips, up to 60mm long, 12mm broad, 8mm thick, ovate-lanceolate acuminate, aristate with bristle up to 5mm long, seldom setiferous, green to grey-green, purplish- green in sun, lines distinct with pronounced small longitudinal transparent areas on upper surface, lower surface heavily mottled or with longitudinal markings, spots occasionally with small hairs, face slightly concave at base, convex-turgid towards upper 1/4, back convex, frequently with second keel or with double row of spines on keel, margins sub-acute, lightly spined, spines white, up to 0.5mm long to 1mm apart. Perianth white, yellowish-green inside, nerves pinkish-brown outside, pinkish inside at tips, tube ascending curved, up to 16mm long, obclavate, bluntly triangular at base, 4.5mm across reducing to 3mm, segments free, regularly stellate, inner lower segments incurved along upper margin. Buds biarcuate, bifid at tips. Flowering Oct-Dec.
Material examined: Cape- 3319 (Worcester): Brandvlei Dam (-CB), Bayer in KG 631/69 (NBG). E. of ridge between Brandvlei Dam wall and Breede
River (-CB), Bayer in KG 20/70 (NBG). NE. of old yacht-club, Brandvlei Dam (-CB), Bayer in KG 141/70. (NBG). NW. of Brandvlei Dam, Pokkraal (-CB), Bayer164 (NBG). Moddergat, 15km S. Brandvlei Dam (-CD), Bayer in KG 226/70 (NBG). Audenburg Peak (-CB), Bayer 1120 (NBG).
Although the precise origin of H. maculata was very vaguely stated, the locality is traceable from the records of G.G. Smith and F.R. Long. A photograph GGS 3912 (fig. 1) is of a plant sent to Smith by Venter as his No.6. It is labelled H. schuldtiana var. maculata v. PoelIn. from Worcester. This matches a photograph by H.G. Fourcade No. 27 of a plant from Long similarly labelled and also GGS 3941 (fig. 2) collected by C. Beukman at the Brandvlei Dam. My own collecting in the Brandvlei Dam area follows a collection by F. J. Stayner in cultivation at the Karoo Garden, which included a form somewhat resembling H. magnifica var. maraisii (V. Poelln.) Bayer. Repeated visits to the area have failed to produce a comparable plant with scabrid retused leaves. This odd plant can be presumed to have been introduced into the collection in error.
 Fig. 1. Haworthia maculata (vPoelln.) Bayer. GGS3912, Venter No.6, Worcester District. |
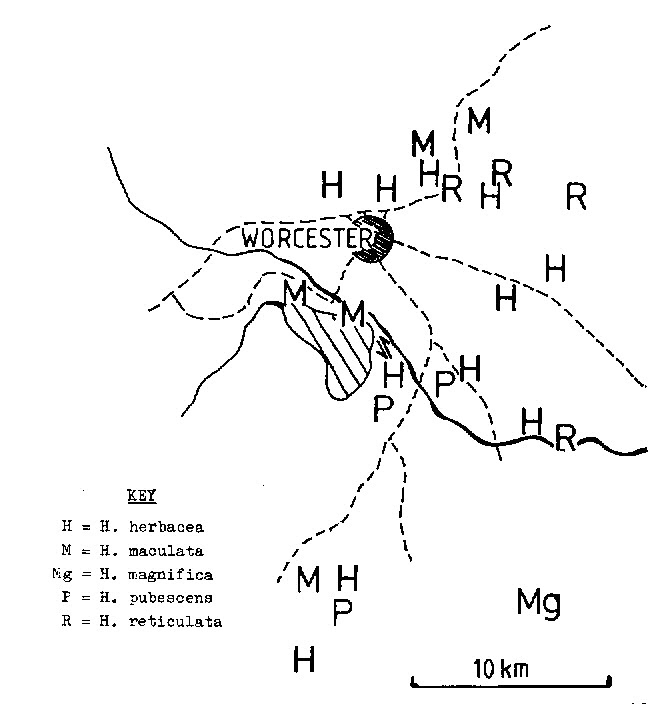
 Fig. 2. Haworthia maculata (vPoelln.) Bayer. GGS3941, C. Beukman, Brandvlei Dam. |
The low hills north of the Brandvlei Dam are in the Witteberg geological series. This is a reddish quartzitic sandstone formation which weathers to acidic low-nutrient sandy soils. The vegetation associated with this formation is Dry Mountain Fynbos. Die Nekkies is the home of the endemic Protea restionifolia and despite the predominance of Fynbos species there are several karoid elements, eg. Drosanthemum hallii, D. bellum, Aloe microstigma, Conophytum ficiforme and Othonna retrofractus. H. maculata is common along the length of these low hills (4-5km) and there is a clinal trend in which plants at the western end have shorter less setate leaves. At the eastern end of the gradient the population is much like that to be expected of an integration of this species with H. herbacea (Mill.) Stear. To the south-east of the Brandvlei Dam wall there is another population (Bayer in KG 141/70) of more robust plants which can clearly be allied to H. submaculata v. PoelIn, which is also a Venter discovery. Although I have placed this element in synonomy under H. herbacea I regard it as the real link with H. maculata. The vegetation at this site is clearly karoid with Pteronia paniculata, Crassula rupestris and Pelargonium alternans on shales underlying the quartzite. The flowers in this population are more typical of H. herbacea, being larger and more beige coloured, in contrast to the easterly population of H. maculata. Here the flowers are distinctly yellow-throated and brown-veined. H. herbacea flowers from Sept-Oct. whereas H. maculata flowers Oct.-Dec. and apparently progressively later from east to west. ( I have also since observed maculate further east at the Cilmor winery where is grows in close proximity to H. pubescens). There is a further population 15km to the south at Moddergat, again in a Fynbos restloid community, which is clearly H. maculata. (Clearly? Rather problematic and this population is as close to maculate as it is to mirabilis as that species becomes confounded with maraisii on the northern fringes of the Riviersonderend Mountains). The only other species in the area apart from H. pumila (L.) Duv. is H. pubescens Bayer.
 Fig. 3. Haworthia maculata (vPoelln.) Bayer. KG 631/69, Brandvlei Dam. |
 Fig. 4. Haworthia maculata (vPoelln.) Bayer 164. NW Brandvlei Dam. |
An almost identical element to H. maculata (which I place with that species) occurs in the Hex River Valley to the north-east of Worcester at Sandhills, and also to the west at a much higher altitude. In this case the plants are in sandstones of the Table Mountain series and the associated vegetation is again Fynbos. Having seen the obvious and dramatic variation in H. habdomadis (v. PoelIn.) as it passes from shales to sandstones, it seems most probable that H. maculata has evolved as an ecotypic derivative of H. herbacea in sandstone habitats. In this case these disjunct populations from north and south of the Breede River valley have most likely evolved independently. At Wolfkloof (Villiersdorp) H. herbacea is similarly modified in a sandstone habitat but with very large pinkish flowers. At the foot of Jonaskop nearby, H. herbacea is unaltered in an unusual karoid shrub community on sandstone.
There is still another dubious population at Buitenstekloof (Bayer in KG 106/70) which I cannot properly place (This was probably the correct intermedia which I may have confounded with reticulata in earlier writing. I have since included this in H. maraisiii var notabilis). This is at a Malmesbury Shale/Dolomite interface, growing with H. reticulata Haw. and H. arachnoidea (L.) Duv. in Mountain Renosterveld. This is approaching the western limit of H. magnifica (maraisii) var. notabilis in a similar habitat north-west of Robertson. The similarities of H. maculata to populations of H. turgida Haw. in sandstone habitats must also be noted. Riley and Majumdar (The Aloineae, 1979) quote my communication to them regarding hybrid swarms quite extensively and I have to retract some of this as misleading. It is obvious that hybridisation of closely related species is a secondary event in the speciation process. The continuity between a whole range of populations must surely be attributable to the speciation process and a reflection of the selection pressures exerted in differing habitats. In the case of H. reticulata and H. herbacea we do have two intimately related species, indistinguishable at one point of a nearly shared distribution range, and visibly distinct and hybridising at another. This phenomenon repeats itself, perhaps less dramatically but over and over again, in Haworthia and other SW. Cape genera.
H. maculata was originally described as a variety of H. magnifica v. Poelln. (i.e. H. schuldtiana v. Poelln. Now H. maraisii) and while the geographic relationship with that species near Robertson has been mentioned, H. magnifica has a complexity of its own which will be dealt with in another article.
(Col. C.L. Scott includes H. maculata in his conglomerate concept of H. asperula Haw. which also includes H. pygmaea v. Poelln., H. pubescens Bayer and all the varieties of H. schuldtiana v. Poelln., a species which I include under H. magnifica. [I later split off the eastern elements as H. maraisii]).
I originally wrote the skeleton of the above for the Haworthia Study Group of New South Wales who printed and distributed the following text:-
Abstract: The status of H. maculata (v. Poelln.) Bayer in relation to its distribution and that of similar species occuring in the Robertson Karoo, is discussed. It is shown that this species is found south of Worcester and in close proximity to H. herbacea (Mill.) Stearn. The previous taxonomic association with H. schuldtiana v. Poelln. was incorrect.
Introduction: The discovery of H. maculata (v. Poelln.) Bayer, is attributed to Major H. Venter, and the locality recorded, as for other species first collected by Major Venter, as in “the neighbourhood of Worcester, Swel!endam, Bredasdorp and Caledon”. The actual locality can be traced by reference to Mr. G.G. Smith’s records and the photographs of Major F. R. Long taken by Dr. H. G. Fourcade. A photograph GGS3912 is of a plant sent to Smith by Venter and labelled H. schuldtiana var. maculata v. Poelln. and this matches Dr. Fourcade’s No. 27 similarly labelled. GGS3941 is a photograph of a plant collected by Mr. C. Beukmann at the Brandvlei Dam south of Worcester, and this is identical to the preceding GGS3912 and No. 27.
A thorough investigation of the Brad. vlei area shows that a. species satisfying the requirements for H. maculata occurs along the length of the low hills which lie between Brandvlei Dam and the Breede River. These hills are a red quartzitic sandstone which is geologically and botanically unique in the Robertson Karoo. A collection of H. maculata made by Mr. F. J. Stayner in this area did include a form somewhat resembling H. schuldtiana, but subsequent collections did not yield another specimen in anyway comparable with H. schuldtlana. A topocline was indicated in which plants at the eastern end of the distribution range had longer and more setate leaves in contrast with plants further west with shorter less setate leaves.
Further investigation revealed the presence of an apparent hybrid complex involving H. maculata and H. herbacea (Mill.) Stear, approximately 1 km. S.E. of the eastern limits of the former species. This appears to be the locality for H. submacula, v. Poelln. collected also by Major Venter, as plants from the locality can be matched with the description for this species. H submaculata has been reduced to synonomy under H. herbacea (Bayer, 1970). Detailed analyses requiring sophisticated rnethods will be needed to properly assess the actual situation at this locality (So I thought at the time – without a sensible impression of the species against a real familiarity of them in the field, no amount of scientific rigour will prove anything.). Superficially it appears that H. maculata and H. herbacea are two distinct taxa either segregating or introgressing (Or existing in close proximity in harmony?). Segregation is favoured as geological factors indicate adaption to different substrate, and botanical factors to different plant communities.
Flower structure is more revealing than the vegetative characters, and at the S.E. locality (KG141/70) the flowers were observed to be more typical of the beige colour in H. herbacea. At the eastern limits of the range of H. maculata (KG631/69) the flowers were distinctly yellow-throated and brownish veined. The flowering time also differed as H. herbacea flowers from October to November, whereas in H. maculata the flowering time extends from October to December, apparently becoming progressively later from east to west.
It could be argued that H. maculata is a subspecies, of H. herbacea. (I do not think anymore that this is a question of objective reality and that argument would generate any kind of accurate decision.) In view of the totality of variation across the range of H. herbacea and H. reticulata, Haw. and in view of other disjunct Haworthia populations in the Robertson Karoo, there is as yet no criterion for treating H. maculata as an infraspecific taxon. This is also bourne out by a plant population at Moddergat (KG226/7O) some 15 km. further south, which undoubtedly belongs to H. maculata. Another population occurs at Lemoenpoort (KG184/70) about 10 km. east of Moddergat, which is of doubtful affinity and totally segregated from H. herbacea growing in the near vicinity (H. pubescens var livida). Still another consideration is the remarkable likeness of certain forms of H. maculata to forms of H. notabilis v. Poelln. at Robertson. However, in this case both flower structure and flowering time indicate that there is less affinity between these species than suggested by vegetative characters.
A population also occurs at Buitenstekloof Langvlei (KG1O6/70) which cannot yet be adequately explained in relation to any of the species so far mentioned us this paper (Later named as H. maculate var intermedia, but since abandoned as a variant of H. maraisii var notabilis)..
Conclusion:
It is concluded that H. maculata Is sufficiently distinct to merit species rank. It is, however, clear that a complex pattern of speciation occurs which justifies a more technical and sophisticated approach than adopted here. The species cannot be associated with H. schuldtiana, and neither does varietal rank adequately express its relationship to H. herbacea. This statement is true especially if the relationship of H. herbacea and H. reticulata is also considered together with other yet poorly understood populations in the Robertson Karoo.
Until a proper basis can be formulated for distinguishing between varietal and species rank in Haworthia there can be little point in attempting to separate these taxa in the genus.
(A formal description followed….)
There is no necessity to compare this species with H. schuldtiana ie. H. maraisii). H. maculata differs from H. herbacea in colour in sunny conditios, is generally more mottled and the marginal spines are always smaller. There are less leaves to the rosette and they are more erect and spreading. The perianth tube is less curved than typical H. herbacea and the flower segments are not as widely flared. Flower colour is different. (Where in H. herbacea the flowers are beige, in maculate they are white – as a generalization. My most recent collection of maculate from the Audensberg has an extraordinarily pink flower). There is no difficulty in separating H. maculata from H. reticulata although plants of H. reticulata from south of Bonnievale may cause difficulty. Here general plant colour, flower colour and proliferation are distinguishing. When H. reticulata is spined, the spines are not as fine as in H. maculata. H. maculata may also be separated from the anomalous H. herbacea/reticulata populations by general facies and as for the separate species.
Separation from H. notabilis on vegetative is difficult and it should be noted that H. notabilis is highly variable across its distribution range at the type locality near Robertson. (The variation increases considerably when one also includes the Klaasvoogds and Agtervink populations to the eat and west respectively.) Flowering time and structure are at variance. Some forms of the H. globosiflora, Smith, and H. nortieri, Smith. complex can be compared vegetatively with H. maculata. (This is a probably a significant statement because as H. maculate occurs in the mountains north of Worcester it is approaching the distribution of H. nortieri north of Ceres.) It should be recognised that a complex species pattern exists in which general facies by which species can be recognised, is the only way of separating them.
Acknowledgement: Gratitude is expressed to Mr. F.J. Stayner, curator of the Karoo Garden, Worcester, for encouragement and assistance during the course of this work. Appreciation of the assistance and interest of Mr. C. Walker and Mr. J. Pilbeam in England, is also expressed.
 Fig. 5. JDV96-34.1 Moddergat |
 Fig. 6. JDV96-34.3 Moddergat |
 Fig. 7. MBB7270.1 Moddergat |
 Fig. 8. MBB7270.2 Moddergat |
 Fig. 9. MBB6815.2 H. maculata. Audensberg. |
 Fig. 10 MBB6815.3 H. maculata. Audensberg. |
 Fig. 11. MBB7526.4 H. maculata. Die Nekkies. |
 Fig. 12. MBB7526.2 H.maculata. Die Nekkies. |
 Fig. 13. MBB7526.5 H.maculata. Die Nekkies |
 Fig. 14. 6815.8 H. maculata. Audensberg. |
 Fig. 15. MBB6815.9. H.aculata. Audensberg. |
 MBB7526.7 H.maculata. Die Nekkies. |