
I will be disappointed if anyone had concluded I had any fixed ideas on the classification of these three species and their relationship. It has a problem which has long been on my mind. What happened recently (Nov.1998) is that I was offered the use of a time-share apartment at Jeffrey’s Bay, near the mouth of the Gamtoos River. I used this opportunity to spend six days in the field testing my hypothesis concerning the species Haworthia cymbiformis, Haworthia cooperi and Haworthia gracilis, and this is what I would like to record. Subsequent to that trip (Mar.1999) I planned and executed an excursion through the Baviaanskloof to Grahamstown and Stutterheim in March 1999, and repeated the exploration in Sept. and Oct, 1999.
Baviaanskloof and environs: 1-Engelandkop, 2-Rooikloof, 3-Moordenaarskloof, 4-Rooihoek, 5-Ripon, 6-Ouplaas, 7-Houtkloof
First I must point out again that there is a problem with plant names in that they do not necessarily reflect a set of objects which can be seen and easily recognised. The naming and typification process is partly to blame. Typification is simply the process of ensuring that a name is based on some substantial recognisable item, preferably a specimen rather than an illustration or photograph. (A specimen is preferred because it lends itself to some other analysis eg. a tissue sample). The type is used to secure and permanently attach a name to for reference purposes. A type specimen and hence a name need not be, and often is not, typical of the taxon which the specimen represents. This does create difficulties particularly when varieties are at issue. In Haworthia there are an inordinate number of populations of uncertain pedigree. This is not because of some obscure evolutionary process, and nor is it just because there is an absence of characters by which to characterise and circumscribe each population. It is what we should expect. Everything is in a state of change and Haworthia is simply following the same unsteady path. I have shown that in Oxalis, in which there are an embarrassment of riches where characters are concerned, that these are of surprising little help in circumscribing the species. I was interested to read this sentence in a book about chameleons, “Without collecting data many chameleons are difficult to identify”. One would expect chameleons to be less difficult to identify than plants. Darwin stated that geographic distribution is the doorway to understanding species, and this is all we have in Haworthia which I can see to make sense.
We simply use ordinary visual observation and sensibility to make judgements about similarities. The less objects we have to examine, the easier it is to assess similarities among them. The problems that then arise are no different from those that develop even with complicated and convoluted discussions and explanations that arise from sophisticated techniques and analysis of characters. In my experience it is very often the direct visual judgement which is used to test the worth of technology rather than the converse. These comments are made because readers seem to think that plant names are as centrally defined as their own unexpressed concept they have of species being groups of things with well-defined limits and readily compartmentalised. Thus they imagine that there is a solution and it is just a question of how it is found. These comments are made because several times in the literature, wild statements have been made about the use of some character or technique or other throwing great light in the darkness. It just does not happen that easily and simply. The more sophisticated the technique, the smaller the sample and the less ‘peer’ review and replication. In statistics it is also pointed out that you do not undertake sophisticated statistical analyses when the figures already show the obvious. Yet how many times do analysts not persevere with test after test until a result does appear significant – itself a matter of probability. Technique may become and end in itself and the purpose for which it is devised lost to view.
There is a nomenclatural code for plant names which regulates about everything except what the binomial stands for. What should be pointed out with some vigour, is that there is a tendency to think that species can be described from single specimens in the paradigm (this is a most useful word because it suggests a pattern, a habit and a model of a time period) of Salm Dyck, Von Poellnitz, G.G. Smith and company. The single plant (and its description) and specimen that is used to typify a name, does not constitute a circumscription of the species. These people described specimens – NOT species. Their work was thus in some measure comparable with stamp collecting. It has little to do with present time description, organisation and understanding of the living variable and changing systems which species are. The different elements in Haworthia named as ‘species’ have been determined by early writers on the basis of vegetative characters such as colour, rosette shape, leaf number and shape, translucence and spination. Nothing much has changed except the scope and extent of the problem. A difficulty with characters is stated in Vavilov’s Law of Homology which states that a set of characters in a taxon will also be present in related taxa. Thus if one finds awned and obtuse rounded leaf tips in the genus, it is possible and indeed probable, that this character set will also occur in lesser ranks. If a set of characters is present in one species, they will possibly and probably occur in related species. This article is to demonstrate the difficulty of classifying plants on this simple basis and in an era when so much more material is there to classify.
It is necessary to discard the name translucens as I have used it in my earlier books. Originally the name really is associated with what is obviously H. herbacea, and I only used it to follow a tradition of misuse. The misfortune of this is that the name gracilis then occupies the nomenclatural hierarchy, but not the same geographic position in my understanding of the species. The consequence is that an outlying variant (Hellspoort, Grahamstown) will become the species name (ie. gracilis) and a name has to be unearthed for the core of the species in the Gamtoos valley viz. H. gracilis var. isabellae. This in my new book replaces H. translucens var. translucens. Similarly H. cooperi var. cooperi is not central to the plot as is the var. pilifera, and I unearthed the latter name in my 1999 revision for this reason, as also the name gordoniana. I opted in 1985 to use the rank ‘subspecies’ under H. translucens for reasons which should be obvious – a weaker geographical relationship of the known variants of the species associated with a broader distributional phenomenon.
This following discussion is done in terms of Darwin’s statement about the importance of geographic distribution, and the evidence I have in my mind for a wide range of species. The essential problem with the species considered in this article is that we have a set of plant populations from Grahamstown, replicated to a degree by a set of populations in the Humansdorp area. In neither area is the relationship in the set well understood, and this does not imply that they can be so understood either. I set out to deal only with recent limited observations in the Humansdorp area but like a stone thrown into a pond, the ripples soon spread.
The link between the Grahamstown and Humansdorp areas is a curious one. It can be seen in the vegetation types of Acocks, which is a broad view of species associations. Valley Bushveld occurs discontinuously from Natal through to Swellendam. Alexandra Thickets are coastal, east of Port Elizabeth. Knysna forest, west of Humansdorp. Dry fynbos dominates the immediate Humansdorp area. Karoid vegetation dominates north of the Groot Winterhoek Mountains. It is at species level that really interesting facts emerge that impact on our analysis of Haworthia. What happens in the genera Aloe, Gasteria, Astroloba etc.? I leave this as a largely unanswered question because unless the reader can respond, he or she is unlikely to appreciate that it may have implications for understanding Haworthia. Astroloba does not occur in the Humansdorp area. There is instead the curious Haworthia pungens. The Baviaanskloof is home to several endemic Gasterias as well as to Aloe pictifolia. Haworthia viscosa is ubiquitous. H. nigra with an otherwise fairly similar distribution is not. The distribution of the genus Encephalartos (cycads) is particularly interesting because of its known antiquity and the similarity of present day species to evidence in the fossil record. Aloe must also have an ancient history as evidenced by presence of species of this genus on Madagascar – a continental-drift pre-history. It would in fact be instructive for would-be authors to read what R.A. Dyer wrote about the classification of cycads, before trotting out any banal aphorism about ‘active evolution’. There is an interesting break (interval) between the cycad species, east of the Sundays and Bushmans River valleys which separate Grahamstown and Humansdorp. This break can be observed in the distributions of other genera and species and evaluated both for compliance and non-compliance. Haworthia has both conditions eg. cymbiformis and pilifera happily cross the interval. Angustifolia and coarctata only just do so. Marumiana, and nigra do so in the inland areas. Attenuata does so. Viscosa is almost ubiquitous. Fasciata does not and neither do sordida, longiana nor xyphiophylla. No species jump the Knysna forest interval and the genus as a whole jumps the Transkei interval northward only via limifolia and koelmanniorum (+mcmurtryi). These east-west breaks are interesting because they thus indicate speciation pressures. They may be helpful in deducing possible or probable relationships in a two-directional way ie. distribution and variation in one set of plant variants provides a tool for the examination of other sets. In the case of H. gracilis, one can argue simply that it complies with the interval, in which case the var. tenera east of Grahamstown is discrete from the Uitenhage, Baviaanskloof look-alikes. Alternatively one can hypothesise non-compliance and that continuity will be found between Grahamstown and Uitenhage. Hellspoort (the site of the var. gracilis) is somewhat supportive of this view, but their is still a wide gap. Perhaps this is only in the known distribution record.
Known collections and observations were the basis of my 1985 classification hypothesis. Many of these were either dry herbarium specimens perhaps supported by photographs of single plants grown in cultivation. This hypothesis for the species in question here was arrived at from specimens from the immediate Grahamstown area and from the lower and eastern Baviaanskloof (Humansdorp) area. For the 1999 revision I have broadened the range of varieties, and relevant to this discussion are:-
| Haworthia cooperi | var. cooperi |
| var. leightonii | |
| var. pilifera | |
| var. gordoniana | |
| and included Scott’s two new species as varieties:- | |
| H. joeyii (= var. dielsiana) | |
| H. venusta (var. venusta) | |
| Haworthia gracilis | var. gracilis |
| var. tenera | |
| var. isabellae | |
| Haworthia cymbiformis | var. cymbiformis |
| var. incurvula | |
| var. transiens |
In this discussion I will largely avoid the issue of both H. bolusii var. blackbeardiana, H. cooperi var. cooperi and H. decipiens although they are closely involved. I will deal primarily with the Humansdorp (Hankey-Patensie); and to lesser degree with the immediate Grahamstown (Plutosvale) area.
I will use the term look-alikes because it is frequently used in the fynbos vegetation to refer to plant species from different genera, which are superficially so similar that close examination and full flowering material is often needed to establish their real identities. In Haworthia this detail is simply not available and thus there are specimens from different species which are geographically and thus probably genetically aeons apart, but which are visually identical.
Grahamstown (Albany)
H. cymbiformis – is well represented and discrete. Yellow-green plants with spreading broader, flatter, ovate leaves. Clump forming plants on steep rocky faces. The var. incurvula – is known from many collections but only one locality (Plutosvale). It has look-alikes in the Patensie area particularly. Its relationship as a variety of cymbiformis really follows G.G. Smith who made the combination on the basis of a continuity which he claims to have seen. I recall (!) that its flowers are more reminiscent of tenera and this article may expose this as a possibility. Pale semi-opaque green, small clump-forming, with ovate, obtuse incurved leaves.
H. cooperi – is well represented by the var. pilifera. Blue-green plants tending to purplish, with incurved thicker, shorter ovate leaves, often spined. Solitary plants withdrawn into the soil. (the var. cooperi is not discussed in this appraisal although the concept presents a substantial problem on which I am preparing a manuscript).
H. gracilis – is known only from Hellspoort north-east of Grahamstown, with a second population of larger more cooperi-like plants from nearer Plutosvale (MBB6603-99, Glen Craig – a Gerhard Marx collection). Gracilis does have look-alikes in the Kirkwood area (Paardepoort, and a greater Humansdorp area). Paler grey-green plants tending to clump, with more leaves. The leaves are incurved narrow attentuate, and with or without spines. My opinion, based on my most recent collection from Hellspoort is that the var. tenera is actually the element concerned and is thus a superfluous name for gracilis. This var. tenera – is very well known from several populations north-east and east of Grahamstown and these include a glabrous population. It has look-alikes in the Uitenhage and Hankey areas of Humansdorp. Grey-green, small solitary or clump-forming, with many narrow, spined incurved leaves.
(Note:- the original description of gracilis seems to be of a moderately spined species, and von Poellnitz concept was apparently drawn from several disparate collections. The conception of the species from an illustration in Desert Plant Life, and indeed my own collection from Hellspoort in early 1970s, is of a moderately spineless plant. The recent collection of mine has forms which are relatively spineless as opposed to forms which are indeed similar to the smaller, densely spined, tenera. Thus we again have the problem where a name is neither central to the geographic element nor typical. The name tenera could be dispensed with and replaced with var. gracilis – this is evident from my discussion).
Humansdorp
H. cymbiformis – represented by the typical variety in the small Baakens River valley and discrete. It may occur north-east of the Baakens River in the Rocklands area of Port Elizabeth. There is also the population at Hell’s Gate north-west of Uitenhage which I am regarding now as the var. transiens (It needs to be re-examined – hastening to add ‘by someone competent to do so’). This var. transiens – is only recently known to be very well represented in the upper Longkloof and in the Baviaanskloof. Its relationship with cymbiformis may not be as close as varietal rank, and the linking population at Hellsgate east of Uitenhage may be misunderstood. Leaves are incurved, pale-green and less opaque than in cymbiformis. Some of the variants resemble plants of the var. incurvula from Plutosvale.
H. cooperi as the var. pilifera – well represented and largely discrete. However, the typical variety with the necrosing-end area seems to have a western limit just outside Port Elizabeth and also near Uitenhage (the furthest west being at Perseverance, JDV90/40). It is then replaced by a variant which is often smaller, having the same blue-green colour darkening to purplish, but the leaf tips remain slender acuminate even in light conditions. This is what I have regarded as the var. gordoniana which in its typical form is not well represented.
H. gracilis as the var. isabellae – is well represented but apparently variable. The leaves tend to be spreading, pale greyish-green and spined. Some plants resemble gracilis from either Hellspoort (less-spined) or from Plutosvale (densely spined – tenera).
General
It is known to me that in the area from Kirkwood, westward to the Little Karoo (north of the Baviaanskloof), there is an interaction between gracilis and decipiens. H. cooperi (not designating any variety) also occurs in that area. There is a problem in the Addo area, where another element (aristata – a separate weak hypothesis) may intrude. It has also been suggested, and this has always been in my own mind, that unicolor (mucronata) from the western Little Karoo may be continuous with either cymbiformis, bolusii or cooperi. It is continuous with arachnoidea, lockwoodii and with decipiens, and I am treating it as a separate issue because there are also other reasons for its exclusion here. The discussion could descend to the point where one says simply that there are not many species in the sub-genus Haworthia.
Results
Grahamstown
I first looked at Hellspoort early in 1970 to collect and form an impression of H. gracilis. This was of a largish plant with leaves exceeding 3cm long, fairly spineless and weakly clustering. I also saw tenera in Plutosvale and at Hunts Drift. The former, small, densely spined and strongly clustering. At a later occasion I collected tenera again in Plutosvale as the conventional spiny element, and close by as a glabrous element of the same size. On that occasion I neither looked for nor saw incurvula or cymbiformis in the area. I revisited Hellspoort with J.D.Venter and G.D.Marx in 1996 and Venter made a re-collection again in 1998 (JDV89/42-117). My 1996 collection (MBB6614-118) reminded me forcefully of Plutosvale tenera, the plants were smaller than my previous collection and also spinier. Venter’s 1998 collection (all slightly different points within Hellspoort) was of the conventional gracilis that I visualise from the Von Poellnitz illustration, and from my early collection. These were quite large plants. During my visit of March 1999, I observed and recorded systematically from the bottom of Plutosvale, to the top (Roff’s Rock). On the north side of the kloof from the Cotswold farmhouse to the middle of the kloof which is Plutosvale, there was tenera both large clusters and relatively solitary plants. I could not locate or identify the site where I had collected the glabrous form before. The area has been degraded by the grazing of goats and it is possible the plants are now gone. From there we explored the south side of Plutosvale as the road climbed out of the kloof. Roffs Rock is very accessible and this no doubt accounts for the large number of herbarium records for incurvula. We found if from virtually the top of Plutosvale, to the lower and eastern point across from where we had observed tenera at our western-most search point. It is possible that the collection G.D.Marx in JDV93/73-93 was from further to the south-west than our starting point at Roffs Rock, at the upper point of Plutosvale. Nevertheless we found the same very pointed leaved plants of that collection, tending to be clump-forming. As we moved eastward, so the plants tended have leaves which were less pointed and more incurved. The colour was yellowish-green rather than the greyer or bluer-green of the Baviaanskloof incurvula look-alikes. The leaves seemed to lack the more swollen upper and end margins that may characterise cymbiformis.
My impression from this repeat visit to the Grahamstown area, is that the elements gracilis and tenera may be better considered as the same taxon. This may lay the basis for a better understanding, or be more compatible, with the range of look-alikes in the west around Humansdorp and Uitenhage.
The incurvula problem is now largely resolved and although it may still be necessary to find other links between incurvula and cymbiformis which Smith claimed and which is not supported by herbarium evidence. To the contrary, the collections of the gracilis–gordoniana plants from Glen Craig and Roffs Rock (viz. -99, -89, -93)), as well as the interplay of species in question in the Baviaanskloof, suggest that incurvula may equally be interpreted as a variant of H. gracilis or even H. cooperi. There is a further possibility based on new observations in respect of H. bolusii var. blackbeardiana. It may be possible to restructure the classification to relate the var. bolusii with H. semiviva; and to consider the element blackbeardiana as a variety of cooperi, (as nomenclaturally correct but geographically weak) as the species, with pilifera and gordoniana as principle variants. Nomenclatural considerations in ‘the strict terms of the code’, might confuse the issue, but I see no reason why reasoned and logical argument could not be used to dismiss names which are confusing because of their geographical connotation.
Humansdorp
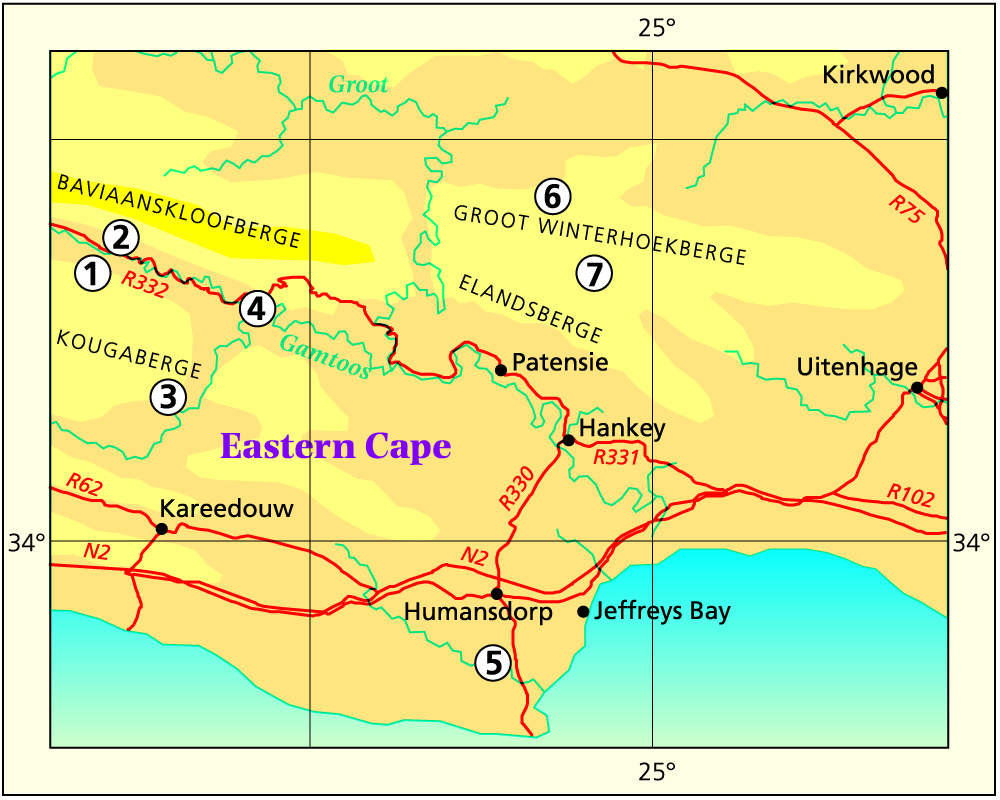
I looked at populations at Jeffrey’s Bay, Zuurbron between Humansdorp and Hankey, two at Draaihoek to the south-west of Patensie, three near Andrieskraal, one further west in the Groot River Poort, and three still further west in the Baviaanskloof, one at Patensie, one at Houtkloof in the Elandriver valley, one at Hankey, one north of Hankey, two south-west of Hankey, and one at the Gamtoos Bridge. Some of these populations were known to me before, and many of them are already represented in the Herbarium. I will also refer to other collections (mostly multiple plants) in my discussion which I have either not seen in the field at all, or in the more distant past. These are collections by credible field botanists and naturalists such as P.V. Bruyns, G.D. Marx, E.J. van Jaarsveld, T Dold and D. Clarke. This thus reviews some 60 populations, but does exclude some significant ones in the Kleinwinterhoek mountains, where other considerations apply.
MBB6792-20 Jeffrey’s Bay. Fynbos. Very short vegetation on level sandstone outcrop. Early bud stage. Small single plants to 20mm diam. Leaves incurved at tips, no marginal spines and bluish-green, darkening to purplish hue (3) – gordoniana–picturata.
MBB6933-21 E. Humansdorp. Grassy fynbos. Rocky sandstone knoll. Small single plants, bluish-green with coarser spination than MBB6793 (2) – gordoniana.
MBB6553-22 and MBB6793-22 Zuurbron. Valley Bushveld. Scrub vegetation on level alluvial stream bank with loose sandstone rocks. Flowering and seeding. Small single plants to 30mm diam. Leaves thicker, leaves incurved at tips, many small closely spaced marginal spines, bluish- green (2) – gordoniana (possibly close to type locality).
MBB6784-23 SW Patensie. Valley bushveld, west sloping conglomerate. Seeding. Small single plants to 30mm diam. Leaves incurved at tips, forms with and without marginal spines, and blue-green (1). Smaller plants of this order were collected by Tony Dold at Drinkwaterskloof in the Baviaanskloof near Geelhoutboskloof – gordoniana.
MBB6786-24 SW Patensie. Valley bushveld, steeper south facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaves incurved at tips, mostly plants with marginal spines, blue-green (2) – gordoniana–isabellae.
MBB6826-25 Rooikloof, mid-Baviaanskloof. Steep conglomerate face, clustering plants – isabellae.
MBB6825-26 Lower Geelhoutboskloof. Cool shaded, steep riverine rock faces. Clustering – transiens–picturata.
MBB6827-27 Rooihoek. East of Geelhoutboskloof. Steep east facing slope. Plants single, greenish, very small – picturata–transiens.
EvJ14680-28 Vetmaakvlakte, S Rooihoek – isabellae (as for a clone of -41, resembling H. aristata).
MBB6789-29 Groot River Poort (Komdomo). Valley bushveld, very steep west-facing shale cliff. Flowering. Small clustering plants to 30mm diam. Leaves with spreading tips, slightly obtuse, no marginal spines, pale-green (5) – transiens–picturata.
MBB6830-30 E Komdomo. Riverine margins. Large clustering plants on steep soil-covered rock- gracilis.
J.N.Reddi in JDV93/86-31 2km NW Andrieskraal, is a collection which may be the same as -30 – gracilis.
EvJ15927-32.1 N. Groot River Poort. Steep south-facing cliffs. Clustering or single. Fairly similar to -29 – transiens–isabellae, the clone 32.2 to decipiens var. minor?
MBB6790-33 Andrieskraal. Valley bushveld, very steep south-facing conglomerate. Flowering and seeding. Small clustering plants to 30mm diam. Leaf tips moderately incurved, slightly obtuse, no marginal spines, pale yellow green (4+Y) – picturata.
MBB6791-34 Andrieskraal. Valley bushveld, steep east-facing conglomerate. Flowering and seeding. Small single plants to 25mm diam. Leaf tips incurved, slightly obtuse, no marginal spines. Pale yellow green (4+y) – picturata.
MBB6930-35.1 E. Andrieskraal, Nuwelande. Steep south facing conglomerate. Small plants down to 16mm diam. As above (4+y). – picturata. JDV90/55-35.2 from nearby westward, is also small plant with very pronounced windows.
MBB6928-36 Nuwelande. Upper steep conglomerate cliff. Plants nearly spineless and the leaves more slender and terete. Greenish (4). These are similar to JDV94/115-37 from the Paul Sauer Dam wall and also from the site for Gasteria glomerata where it was collected by E.van Jaarsveld (EvJ11076 in JDV90/118-38.1&2). A plant from that collection (-38.3) also resembles the Ripon plants (MBB6932-39) which have broader more deltoid, and more spinose leaves – isabellae.
J.G.Marx298 in JDV94/30-40, also Paul Sauer Dam, include both spined and spineless plants which also appear to be (4) – isabellae.
MBB6799-41.1 also (-9&-10) NW Patensie. Valley bushveld. South facing conglomerate slope. Seeding. More robust single plants to 45mm diam. Leaf tips incurved, with and without marginal spines. Pale green and blue-green (1-3) – gracilis–isabellae. One clone in this quite variable collection, strongly resembled a large specimen of H. aristata (-125,-142,-143).
MBB6801-42 SW Hankey. Early flowering. Valley bushveld. Vertical conglomerate. Small clustering plants to 20mm diam. Leaf tips incurved, with marginal spines. Pale blue-green (2-3) – isabellae.
MBB6802-43 SW Hankey. Valley bushveld. Steep east facing conglomerate. Early flowering. Plants mostly single to 30mm diam. Leaf tips tending to spread, with marginal spines. Pale blue-green (2-3) – isabellae.
PVB7128-44 Holriver, far S Patensie. Steep north-facing cliffs. Similar to above – isabellae.
D.Clark1050-45 Kouenek, Geelhoutboskloof. No detail, similar to above. – isabellae.
MBB6773-46 N Kareedouw. Fynbos. Conglomerate. South facing. Flowering late Dec. Plants single small, leaves incurved, very spiny and spines on leaf surfaces too. Blue-green (2) – isabellae.
MBB6771-47 Moordenaarskloof (N. Kareedouw). Steep south facing slope. Plants in small clusters or single. Relatively spineless. Leaves incurving. Green (4) – picturata.
MBB6805-48 NE Hankey. Valley bushveld. Gradual west facing slope. Seeding. Plants single to 50mm diam., leaf tips incurved, with and without marginal spines. Blue-green and pale-green and (1-4) – gracilis–isabellae.
MBB6804-49 N Hankey. Valley bushveld. Dense vegetation on alluvial stream bank. Seeding. Plants single to 50mm diam., leaf tips incurved, without marginal spines. Pale-green and blue-green (1-4) – gracilis–isabellae.
JDV97/3-50 E Hankey. Valley Bushveld. Plants similar to above – (1-2) gracilis–isabellae.
MBB6808-51.1 Gamtoos Bridge. Valley bushveld. Steep west facing conglomerate. Seeding. Clustering plants to 50mm diam. Leaf tips spreading, with marginal spines. Blue-green (2) – isabellae. There are very similar plants at Longmore, east of Loerie (MBB6555-51.2).
MBB6798-52 Houtkloof (Upper Elandsriver valley). Fynbos. Rocky valley in sandstones. Neither flower nor seed (flowered in cultivation in Jan.). Plants single to 35mm diam. Leaf tips incurved, very spiny. Blue-green (2) – isabellae.
C.Marais in JDV96/95-53 Forest Glade and JDV92/136-54 Oaklands, both Elandsriver, are similar but more coarsely spined and paler green (4) – isabellae.
MBB1404a in JDV86/13-55 is an old collection of a tenera-like plant from the lower Elandriver valley (Groendal Dam) – cf. tenera.
D.M.Cumming6831 is from nearby and is more typically isabellae-like).
PVB7040 in JDV97/8-57 and PVB7944 in JDV97/7-58 Ouplaas (E Cockscomb) are very similar to the above from the Elandsriver, but the plants are more robust, with much wider and more obtuse leaves. Particularly interesting because similar plants also occur in the Kleinwinterhoek eg JDV93/41-59 N Campherpoort, JDV87/180-60 S Campherpoort, JDV91/136-61 N Wolvefontein, JDV92/140-62.1 and JDV94/45-62.2 both Wolven (all decipiens var. minor). There is also the collection from the northern Groot River Poort (EvJ15927 – 32.2) linking these, and perhaps also the Cockscomb and Elandsriver plants (ie. isabellae) with transiens.
JDV97/6-63 W Braamrivier. Dry Fynbos. Steep W. facing cliff. Clump-forming, short obtuse incurving leaves. Pale green (5) – transiens. EvJ15342-64 Dieprivier, NE Kareedouw; EvJ17548-65 Horee, Saptou, SE Uniondale; PVB7077 in JDV97/5-66.1 Oshoek, E of Uniondale; MBB6729-66.2 S Uniondale, and Reddi in JDV93/54-67 Kabeljouwsriver, near Jeffrey’s Bay (a doubtful collection), are also this element – transiens.
The latter is similar to -29 Groot River Poort (transiens–picturata), ie. small clustering plants to 30mm diam. Leaves with spreading tips, obtuse, no or very few marginal spines, pale-green. I could not locate this population and it would prove that gracilis, transiens and cooperi co-exist. E. Aslander1247-68 made a collection from the place reported by Reddi, and these plants are unquestionably gordoniana-like.
PVB7093 in JDV97/1-69 Skrikrivier, north of -63 is similar to that collection, but leaves more elongate acuminate and some lightly spined. Flowered late Jan. Very reminiscent of gracilis var. gracilis and particularly a brighter green variant from Paardepoort (MBB6600-70, var. viridis in the new revision) in the Kleinwinterhoek mountains to the north-east. PVB5402 in JDV97/20-71.1 Palmietrivier and MBB6589-71.2 Dorschfontein, both E Steytlerville, are similar to this latter collection but better related to decipiens var. minor. A problem arises here in trying to limit the scope of the paper because there are similar collections from north of Glenconnor (JDV91/17-72.1), and south of Lake Mentz (JDV91/116-72.2, which grows with gordoniana JDV91/115-73).
MBB6810-74 (also -11&-12) Joubertina. Fynbos. South facing, rocky sandstone slope. Neither flower nor seed. Plants single to 50mm diam. Leaf tips incurved, without marginal spines. Blue-green (1) – gordoniana.
MBB6811 & JDV90/80-75 (also -13) Uniondale Pass. Renosterveld. Rocky defile, steep sandstone. Early bud stage. Plants single to 25mm diam. Leaf tips incurving, with and without marginal spines. Blue-green (1) – gordoniana. PVB7079-76 Saptou, (Upper Longkloof), PVB7062-77 Redcliffe (NE Willowmore), JDV91/80-78 Engelandsekloof (Baviaanskloof), JDV94/95-79 Nuwekloof (W. Baviaanskloof), are all in this class. A collection G Marx194 in JDV91/81-80 from Apieskloof (Baviaanskloof) is unusually pale green but otherwise also seems to compare with these gordoniana-like plants).
In completing this review of collections I would like to mention five which seem to touch on this issue, but which are even more relevant to the H. cooperi var. cooperi and H. bolusii var. blackbeardiana issue. These are G Marx in JDV91/14-145 De Plaat (NW Kirkwood), PVB5002 in JDV92/33-147 Kaboega Gorge (Suurberg), JDV96/89-116 Gladhurst (S Adelaide), J.G.Marx in JDV96/4-98 and MBB6603-99 NE Grahamstown (Glen Craig), and JDV93/73-93 upper Plutosvale. These are all plants with elongate acuminate leaves about as wide as thick, and almost or completely without spines. They could be confused with H. cooperi var. leightonii, or with clones of H. bolusii var. blackbeardiana – and of course with H. gracilis var. gracilis.
Discussion
In trying to circumscribe each collection I have been aware of my deficient descriptive skills. Can one rationalise such similar things one observes in writing in a way and which can lead others to identifications?. I have elsewhere pointed out major weaknesses in the capacity of persons to compare what is written with what is seen, and also to compare plants and illustrations. Apart from the problem of variation in the populations, there is a problem of different habitat conditions (light, soil, temperature and moisture) which will cause phenotypic differences. It should be possible to define these if the plants could all be grown under the same conditions. However, local variation and sample size also becomes a problem. Colour must be critical. There are four main classes in this set of species under discussion – the opaque yellow-green of cymbiformis, the darker blue-green of cooperi which can develop reddish veins and a purplish colour under stress, the mid pale (or light) grey-green of translucens and the subdued pale green of transiens. Under stress the latter becomes very pale. In the descriptions above I have categorised colour on a scale of 1-5 from blue-green through to pale green, and added Y to indicate a more opaque yellow colour. In my book I recognise a variety (ie. MBB6791-34 picturata from Andrieskraal) in which the transition from opacity to translucence is abrupt. This collecting trip does not strongly support such a geographic entity although their may be populations in that area which do. Certainly in the var. transiens, some plants are also very conspicuously patterned. Flowering time in early summer for all these collections appears to be very much the same. Curiously the Jeffrey’s Bay collection was still at bud stage when most of the other collections were seeding already. When I visited this population six weeks earlier in the previous year, the plants were already in flower.
The off-setting, clustering character is also worth commenting on. As in the case of H. turgida and H. retusa (remembering also an expressed contention of mine that the latter can be regarded as a variant of the former), clustering is usually associated with very steep rocky situations as opposed to plants on level sites being solitary. In this particular study the slopes varied from level through moderate to vertical. Clustering was most pronounced on vertical sites and at Andrieskraal where vertical and sloping sites were nearly adjacent, the plants were clustering on the vertical and solitary on the sloping. Clustering may be accompanied by the capacity of the plant stems to elongate. It is significant that populations around Hankey and Patensie which seemed to be var. gracilis, were invariably on moderate sloping or level ground (NOTE. I have previously always identified these collections as gordoniana), and using the name gracilis now is derived from the product of the papers in the compendium. My later papers suggest that transferring these elements to H. cooperi is worth consideration).
The above collections confirm my observation that Haworthia in the Baviaanskloof are very difficult to classify and circumscribe. It does seem on the basis of collections by Bruyns, Van Jaarsveld, and Venter that transiens is a very substantial element to the west, which may not have a strong connection with cymbiformis. Gordoniana is apparently a much stronger element than I had previously supposed and I am comfortable with the decision to now recognise it as a valid variety of cooperi. The above collections in the Longkloof, and some other recorded collections, seem to show convincingly that it effectively co-occurs with transiens. In the area just west of Patensie, three collections described above seem to indicate that gordoniana intergrades directly with transiens and this transition gives rise to plants which resemble incurvula and which I identify as H. gracilis var. picturata. Similarly south-west of Patensie gordoniana (-24) seems to alter to isabellae. South west of Hankey this latter variety (-42) resembles tenera. The resemblance to tenera is also apparent in the Groendal area of Uitenhage. The Elandsriver collection (-52), and a similar one by Clifton Marais in the same valley (-53), appear to be intermediate in appearance between tenera and isabellae. Isabellae as a separate entity is also represented by the Gamtoos Bridge collection (-51), and an old collection by W.R. Branch from the Krom River estuary at Ripon (-39).
A major difficulty is the Jeffrey’s Bay population (-20). These small solitary plants seem, by virtue of their growth habit and the near co-occurrence with isabellae (perhaps transiens too, if Reddi’s collection -31, was confirmed), to be the local equivalent of cooperi var. pilifera. However, if they had been observed at Grahamstown, they would be taken to be gracilis. In cultivation they have come to bear a stronger resemblance to the -47 which is closer to picturata than to gordoniana. The Zuurbron population (-22) passes for the equivalent of pilifera with some difficulty, and this does transpose to isabellae via Draaihoek, SW of Patensie (-24). The gracilis-like plants around Hankey (-48,-49,-50) and Patensie (-41) too, often present an isabellae facies. Thus we are faced with the possibility of a strange inversion. The upper Longkloof has cooperi as the var. gordoniana, but in the lower Longkloof similar plants may be derived from gracilis. A similar problem is presented by the similarity of gordoniana-like plants to H. bolusii var. blackbeardiana at several different places in the wider Eastern Cape.
The collection -46 is particularly interesting (I am indebted to Ernst van Jaarsveld for its discovery). This is a very small isabellae which has the same colour and (spininess) hairiness of the leaf surfaces as has Scott’s venusta. This may be very significant in providing supporting for an hypothesis that prior to inundation of the continental shelf (12-18000yrs ago) there was less of a vegetation interval east and west of the Sundays and Bushmans Rivers. I have suggested that this is reflected in the relationship and distribution of H. coarctata and H. reinwardtii, and will suggest elsewhere that H. fasciata may also be evidence of such a situation. However, this is very speculative. The surface spination is also evident in some collections from north and south-west of Jansenville (-129,-130,-131,-132 and -133 which I have identified with either decipiens vars. minor or pringlei), although the spination is sparse.
Conclusions
These specific observations suggest that my classification hypothesis (the classification as presented in my two handbooks and in my recent revision) is a sound communication medium for the two areas in question, without implying thereby that it cannot be improved. What is problematic is that in the greater Baviaanskloof area, the three elements never seem to occur together. I repeat what I have written in respect of H. cooperi and H. bolusii var. blackbeardiana, and in my handbooks and revision. It is also stated indirectly in the preamble to this paper. “The essence of my classification is that of co-occurrence and to examine its merit it can be tested on that basis. Botanists would probably prefer the use of the terms sympatric and allopatric, but I think technical language does often just strain goodwill”. In this particular article, the key question would have been:-
“Do three elements we can identify as separate living systems representing the species cooperi, cymbiformis and gracilis grow in the same close geographic proximity?”.
Evidence available to me and in the herbarium record, is that they do not. There are a few known instances where two of these species are in such proximity – but not all three. What is curious is that it is rare for any two species of the subgenus Haworthia to be in immediate proximity to the extent that they share any specific habitat.
Observations seem to confirm my opinion that there is a connection between the three species in their occurrence east and west of the Sundays and Bushmans Rivers. What does concern me is the relation between cymbiformis and transiens, and the implications these observations have for incurvula, gracilis and cymbiformis around Grahamstown. The set of plants around Grahamstown favours cooperi, cymbiformis and tenera, while the mirror set of the Humansdorp area, favours gordoniana, transiens and isabellae. The relationship of incurvula needs to be explored. In the eventual integration of all these collections, sight should not be lost of the complex situation around Fort Beaufort, where the same set of species and variants seems to be present. Nor should it be forgotten that in the Kleinwinterhoek mountains H. decipiens becomes involved. The situation north of the Groot River is complex and there are substantial records from the Perdepoort and from Willowmore and Steytlerville which indicate a close interaction between decipiens and gracilis as well as gordoniana.
There are a good many other collections from the upper Baviaanskloof, but mostly these are represented only by herbarium records and I would like to see this material in its living state.
A final thrust of this article is to emphasise that Haworthia is not going to be understood by botanists or enthusiasts sitting on some other continent. This is particularly if they depend on the convolutions of the Code of Nomenclature, literature and limited collections for their information and the opinions they feel compelled to express. Less so if they do not and cannot, read and write the english language sufficiently well to communicate properly. Still less so if they have no knowledge and feel of South African geography and vegetation, and are operating in an outdated paradigm where specimens represent entire species. Explanations which only account for a few plants in cultivation are not going to be very satisfactory. The fact must be faced that classification is difficult and more information makes it more difficult. This is not a problem peculiar to Haworthia. It only seems so because the genus has received so much attention. The question now is:
“How is this information recorded and presented in a way that can be accessed, understood and appreciated?”.
My conclusion is that we now need to resort to computerisation and digital imagery, recognising that dealing with single plants is not the same as dealing with populations. It is not easy to assess populations objectively as the sample sizes become unmanageable. We should progressively assess and re-assess populations and build up good herbarium documentation. In this way a proper revisionary process is set in motion, which limits Micky Mouse decisions about hypotheses, classification and nomenclature.

1 a. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay. 
1 b. MBB6792 H. cooperi var. gordoniana to H. gracilis var. picturata, Jeffrey’s Bay. 
2 a. MBB6933 H. cooperi var. gordoniana, east of Humansdorp. 
2 b. MBB6933 H. cooperi var. gordoniana, east of Humansdorp. 
2 c. MBB6933 H. cooperi var. gordoniana, east of Humansdorp. 
3 a. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron. 
3 b. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron. 
3 c. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron. 
3 d. MBB6553 & MBB6793 H. cooperi var. gordoniana, type locality, Zuurbron. 
4 a. MBB6784 H. cooperi var. gordoniana, southwest of Patensie. 
4 b. MBB6784 H. cooperi var. gordoniana, southwest of Patensie. 
5 a. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie. 
5 b. MBB6786 H. cooperi var. gordoniana to H. gracilis var. isabellae, southwest of Patensie. 
6 a. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof. 
6 b. MBB6826 H. gracilis var. isabellae, a tiny form, Rooikloof, mid-Baviaanskloof. 
7. MBB6825 H. cymbiformis var. transiens to H. gracilis var. picturata, lower Geelhoutboskloof. 
8. MBB6827 H. gracilis var. picturata to H. cymbiformis var. transiens, east of Geelhoutboskloof. 
9. EvJ14680 H. gracilis var. isabellae, Vetmaakvlakte, south of Rooihoek. 
10 a. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
10 b. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
10 c. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
10 d. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
10 e. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
10 f. MBB6789 H. cymbiformis var. transiens to H. gracilis var. picturata, Groot River Poort (Komdomo). 
11 a. MBB6830 H. gracilis var. gracilis, east of Komdomo. 
11 b. MBB6830 H. gracilis var. gracilis, east of Komdomo. 
11 c. MBB6830 H. gracilis var. gracilis, east of Komdomo. 
12. JDV93/86 H. gracilis var. gracilis, 2km northwest of Andrieskraal. 
13 a. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
13 b. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
13 c. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
13 d. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
13 e. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
13 f. EvJ15927 H. cymbiformis var. transiens to H. gracilis var. isabellae, northern Groot River Poort. 
14 a. MBB6790 H. gracilis var. picturata, Andreskraal. 
14 b. MBB6790 H. gracilis var. picturata, Andreskraal. 
14 c. MBB6790 H. gracilis var. picturata, Andreskraal. 
15 a. MBB6791 H. gracilis var. picturata, Andrieskraal. 
15 b. MBB6791 H. gracilis var. picturata, Andrieskraal. 
15 c. MBB6791 H. gracilis var. picturata, Andrieskraal. 
15 d. MBB6791 H. gracilis var. picturata, Andrieskraal. 
15 e. MBB6791 H. gracilis var. picturata, Andrieskraal. 
16. 1 a. MBB6930 H. gracilis var. picturata, east of Andrieskraal. 
16. 1 b. MBB6930 H. gracilis var. picturata, east of Andrieskraal. 
16. 1 c. MBB6930 H. gracilis var. picturata, east of Andrieskraal. 
16. 1 d. MBB6930 H. gracilis var. picturata, east of Andrieskraal. 
16. 2 JDV90/55 H. gracilis var. picturata, also east of Andrieskraal. 
17 a. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal. 
17 b. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal. 
17 c. MBB6928 H. gracilis var. isabellae, Nuwelande, east of Andrieskraal. 
18. JDV94/115 H. gracilis var. isabellae, Paul Sauer Dam Wall. 
19.1. EvJ13809 H. gracilis var. isabellae, upper Paul Sauer Dam. 
19.2. EvJ11076 in JDV90/118 H. gracilis var. isabellae (smooth), upper Paul Sauer Dam. 
19.3. EvJ11076 in JDV90/118 H. gracilis var. isabellae (= MBB6932, Ripon), upper Paul Sauer Dam. 
20. MBB6932 H. gracilis var. isabellae, Ripon, Krom River. 
21 a. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam. 
21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam. 
22. 1 a. MBB6799 H. gracilis var. isabellae, northwest of Patensie. 
21 b. J.G.Marx298 in JDV94/30 H. gracilis var. isabellae, Paul Sauer Dam. 
22. 1 c. MBB6799 H. gracilis var. isabellae, northwest of Patensie. 
22.2. MBB6799 H. gracilis var. gracilis to H. gracilis var. isabellae, northwest of Patensie – a clone resembling H. aristata. 
23 a. MBB6801 H. gracilis var. isabellae, southwest of Hankey. 
23 b. MBB6801 H. gracilis var. isabellae, southwest of Hankey. 
24 a. MBB6802 h. gracilis var. isabellae, southwest of Hankey. 
24 b. MBB6802 h. gracilis var. isabellae, southwest of Hankey. 
25 a. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie. 
25 b. PVB7128 H. gracilis var. isabellae, Holrivier, far south of Patensie. 
26. D.Clark1050 H. gracilis var. isabellae, Kouenek, Geelhoutboskloof. 
27 a. MBB6773 H. gracilis var. isabellae, north of Kareedouw. 
27 b. MBB6773 H. gracilis var. isabellae, north of Kareedouw. 
28 a. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw. 
28 b. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw. 
28 c. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw. 
28 d. MBB6771 H. gracilis var. picturata, Moordenaarskloof, north of Kareedouw. 
29 a. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey. 
29 b. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey. 
29 c. MBB6805 H. gacilis var. gracilis to H. gracilis var. isabellae, northeast of Hankey. 
30 a. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey. 
30 b. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey. 
30 c. MBB6804 H. gracilis var. gracilis to H. gracilis var isabellae, north of Hankey. 
31 a. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey. 
31 b. JDV97/3 H. gracilis var. gracilis to H. gracilis var. isabellae, east of Hankey. 
32.1 a. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge. 
32.1 b. MBB6808 H. gracilis var. isabellae, Gamtoos Bridge. 
32.2 a. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie. 
32.2 b. W.R.Branch in MBB6555 H. gracilis var. isabellae, Longmore Forest, east of Loerie. 
33 a. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley. 
33 b. MBB6798 H. gracilis var. isabellae, Houtkloof, Upper Elandsriver valley. 
34 a. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver. 
34 b. JDV96/95 H. gracilis var. isabellae, Forest Glade, Elandsriver. 
35 a. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver. 
35 b. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver. 
35 c. JDV92/136 H. gracilis var. isabellae, Oaklands, Elandsriver. 
36 a. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage. 
36 b. MBB1404a in JDV86/13 H. gracilis var. tenera, Groendal Dam, west of Uitenhage. 
37. D.M.Cumming6831 H. gracilils var. isabellae, south of Groendal Dam. 
38 a. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb. 
38 b. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb. 
38 c. PVB7040 in JDV97/8 H. gracilis var. isabellae, Ouplaas, east of Cockscomb. 
39. PVB7043 in JDV97/7 H. gracilis var. isabellae, Ouplaas, east of Cockscomb. 
40 a. JDV93/41 H. decipiens var. minor, north of Campherpoort. 
40 b. JDV93/41 H. decipiens var. minor, north of Campherpoort. 
41. JDV87/180 H. decipiens var. minor, south of Campherpoort. 
42 a. JDV91/136 H. decipiens var. minor, north of Wolvefontein. 
42 b. JDV91/136 H. decipiens var. minor, north of Wolvefontein. 
43.1. JDV92/140 H. decipiens var. minor, Wolven near Wolvefontein. 
43.2. JDV94/45 H. decipiens var. minor, Wolven near Wolvefontein. 
44 a. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier. 
44 b. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier. 
44 c. JDV97/6 H. cymbiformis var. transiens, west of Braamrivier. 
45 a. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw. 
45 b. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw. 
45 c. EvJ5342 H. cymbiformis var. transiens, Dieprivier, northeast of Kareedouw. 
46. EvJ15748 H. cymbiformis var. transiens, Horee southeast of Uniondale. 
47.1 a. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale. 
47.1 b. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale. 
47.1 c. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale. 
47.1 d. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale. 
47.1 e. PVB7077 in JDV97/5 H. cymbiformis var. transiens, Oshoek, east of Uniondale. 
47.2. MBB6729 H. cymbiformis var. transiens, De Vlugt, Prince Alfred’s Pass, south of Uniondale. 
48. J.N.Reddi in JDV93/54 H. cymbiformis var. transiens, Kabeljouwsriver near Jeffrey’s Bay. 
49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay. 
49. E.Aslander1247 H. cooperi var. gordoniana, Kabeljouwsriver near Jeffrey’s Bay. 
50 b. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c]. 
50 c. PVB7093 in JDV97/1 H. cymbiformis var. transiens to H. gracilis var. viridis, Skrikrivier, north of JDV97/6 [44 a-c]. 
51 a. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains. 
51 b. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains. 
51 c. MBB6600 H. gracilis var. viridis, Paardepoort, Kleinwinterhoek mountains. 
52.1 a. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville. 
52.1 b. PVB5402 in JDV97/20 H. decipiens var. minor, Palmietrivier, east of Steytlerville. 
52.2 a. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville. 
52.2 b. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville. 
52.2 c. MBB6589 H. dicipiens var. minor, Dorschfontein, east of Steytlerville. 
53.1. JDV91/17 H. gracilis var. viridis, north of Glen Connor. 
53.2. JDV91/116 H. graciis var. viridis, south of Lake Mentz. 
54 a. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort. 
54 b. JDV91/115 H. aristata to H. cooperi var. gordoniana, Sundays River Poort. 
55 a. MBB6810 H. cooperi var. gordoniana, Joubertina. 
55 b. MBB6810 H. cooperi var. gordoniana, Joubertina. 
56 a. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass. 
56 b. MBB6811 & JDV90/80 H. cooperi var. gordoniana, Uniondale Pass. 
57. PVB7079 H. cooperi var. gordoniana, Saptou upper Longkloof. 
58. PVB7062 H. cooperi var. gordoniana, Redcliffe northeast of Willowmore. 
59. JDV91/80 H. cooperi var. gordoniana, Engelandkop, Baviaanskloof. 
60 a. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof. 
60 b. JDV94/95 H. cooperi var. gordoniana, Nuwekloof, western Baviaanskloof. 
61 a. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof. 
61 b. J.G.Marx194 in JDV91/81 H. cooperi var. gordoniana, Apieskloof, east of Geelhoutboskloof, Baviaanskloof.