
Many years of experience have left me very skeptical of botany and botanical science, and particularly of writers who masquerade as taxonomists. This is reflected in much of my writing and in these essays. One of the difficulties I have had is in trying to understand why botanists tend to regard species as quantum units which are distinguishable from one another unless some taxonomists has made some statement to suggest that they are not. Non-botanists are further beyond the pale. This brief essay is to demonstrate that I have not found vaunted sophistication and technology of much assistance in better understanding the relationship of Haworthia populations to one another. Often references to such technology are used as subterfuge by authors who should not be engaging themselves in classification at all.
Early in 1970 Dr David Cutler (Royal Botanic Gardens, Kew) was kind enough to have made many photographs of the leaf surfaces of a large number of Haworthia specimens representing a wide range of seemingly interconnecting populations. See the attached list for the plants examined. I ended up with about 350 of these photographs which I have been unable to explain. Looking at them as one would look at the plants themselves, I am again and again struck by the fact that in the end, single specimens were taken to represent populations. This is despite a specific request to have at least 10 leaves photographed from at least one population. The photographs were taken at various magnifications, powers and angles. Most significant would probably have been 100X, but only about 10 such pictures were taken. The most were at 300X magnification and this article is illustrated with these, excepting the final fig.24 which is 100X).
I could not, and can still not, find any pattern which helps me to resolve the relationships any better than explained in many of my publications or in my revision of 1999. No population can be described from a single plant, and in Haworthia no meaningful description can be written to exclude plants from other populations – such is the nature of the continuities that exist between populations. To expect that leaf surfaces (and flowers and seed too, as has so often been implied by my critics) will provide characters to resolve these continuities into recognisable discrete entities, is actually very far-fetched. Suggesting that it will, borders on the fraud. The most I would expect is that leaf surface morphology as depicted in an ESM photograph, may be very useful for distinguishing between, say, H. viscosa and H. pungens, where nothing is actually needed; but it is not going to resolve the problems I described, say, in H. rossouwii and the complexities of its relationships with as many as seven other elements. These elements can be seen to be discrete at some point in their distribution ranges, but not everywhere. I mention H. viscosa and H. pungens particularly because there is a writer who has already proclaimed the latter to be a re-description of H. beanii, despite the very obvious differences that are visible to any eye. They are not the same thing, and the latter is in fact a variant of H. viscosa. These inaccurate diagnoses and ridiculous statements are all too common and P. Halda made a similar error. He described a new variety of H. viscosa from the Baviaanskloof, not far from the localities for H. beanii and its var. minor (both G.G.Smith). However, he also went so far as to also reduce H. nigra to the rank of subspecies under H. viscosa. These are major issues which are difficult to refute in the absence of incontrovertible evidence, despite the fact that they are so easily resolved by reference to the field situation and gross visual examination. One can separate H. viscosa (H. beanii) and H. pungens in the dark simply by feel.
In the subgenus Haworthia, the problems are wholly of a different magnitude. The various species cannot be differentiated by any method and never will be – until the resolution comes to pass of its own in geological time.
Looking at some of the SEM photographs:-
Fig.1 is KG226/70 H. maculata from Moddergat, south of Worcester. In terms of the photographs I have, it is very comparable with fig.2 KG48/70, which is H. maraisii from Stormsvlei, and it may also be comparable with fig.3 KG20/70 which is also H. maculata but from the Brandvlei Dam south of Worcester. But then I am confronted with four other photographs for this species, viz. figs.4 and 5 also Moddergat, and figs.6 and 7 both Brandvlei. One can easily agree on some or other common feature and perhaps decide that we are dealing with one species. This is not the case.
We can take fig.8, which is also KG20/70 H. maculata from Brandvlei, and juxtapose it with fig.9 KG163/71 H. emelyae var. multifolia from Springfontein in the Little Karoo – very comparable. Figs.10 KG178/70 H. reticulata and Fig.11 KG 179/70 H. notabilis are both from Wolfkloof north of Robertson. They are barely distinguishable and especially so in the context of the degree of difference between photographs of the same specimens at different power and angle.
Figs.12 and 13 KG36/70 are H. heidelbergensis from Rooivlei north of Bredasdorp. This is a problem population because it may have (it does have) ties with fig.14 KG318/71 H. rossouwii (was H. serrata), Oudekraal, Heidelberg, and with fig.15 KG709/60 H. mirabilis var. sublineata, south of Bredasdorp. The latter must be compared with fig.16 which is the latter element again, but from north of Bredasdorp. Two other figures are fig.17 and fig.18, KG81/71 H. heidelbergensis, from Matjestoon south of Heidelberg and KG210/71 H. heidelbergensis from just east of Heidelberg. There is very little evidence of discrete pattern.
If one attempts to formulate pattern from the above figures, then look at fig.19 KG134/70 H. mirabilis var. triebneriana (the forma diversifolia) from east of McGregor, or fig.20 KG213/70 H. maraisii from the sandstones south of McGregor. Then I include fig.21 KG35/70 H. maraisii from north of Bredasdorp and compare with fig.22 which is H. mucronata var. inconfluens from west of Ladismith.
I emphasise that these are just single samples from an extremely limited range of populations and I cannot imagine that anyone can afford the time and the expense of analysis of enough material, to arrive at any definite and better conclusion. It is a simple phantasm to suggest that technology is going to resolve a conceptual problem. Where aspiring writers are unable on macromorphological grounds to resolve issues as obvious as that of H. pungens and H. viscosa or H. viscosa and H. nigra, technology and sophistication are potential weapons of folly.
From all the pictures, it emerges that H. maraisii and H. mirabilis may have a commonality in the raised nipple at the centre of the surface cells. However this is absent in fig.23 KG400/61 H. maraisii (a variant – naturally) from Dublin west of Robertson, while it is present in H. arachnoidea and specifically fig.22. Fig.23 represents quite a major departure from most of the photographs and it will attempt someone to suggest this must be a different species. This will be ridiculous. My species concept and definition is that geographical distribution is the major criterion for the determination of the species. Thus the plant from which fig.23 is derived is firstly seen as from a single population which is just a geographical part of an extended complex. Without any strain on logic or reason, its characteristics can be taken to extend from, and also contribute to, that complex. Thus if leaf surfaces are to be studied in pursuing understanding of the relationships in Haworthia, a good approach would be to first take such a single complex and take an overview. The photographs suggest that 100X would have been a good magnification at which to do this (see fig.24 H. notabilis and compare with the same in fig.11).
What needs to be done is for my revision and explanation of Haworthia to be taken as a hypothesis and tested as such. Even an article of this kind requires this basic hypothesis. The presentation of alternate hypotheses and nomenclatural solutions based on less material, inadequate conceptualization and poor intellectual preparedness is a disaster for the collector and enthusiast. It is also a disaster for the botanist and scientist who fail to understand, as so many do, that the authority of a classification is not just a question of acceptance or observance of a nomenclatural code and the product of anyone who is enthusiastic enough to produce an identification key and a classification. There is adequatio involved in the case of both the taxonomist and the acceptance of the product.

Fig. 1. MBB in KG226-70 H. maculata. Moddergat, S Worcester. 
Fig. 2. MBB in KG48-70 H. maraisiivar maraisii. Stormsvlei. 
Fig. 3. MBB in KG20-70H. maculata. Brandvlei. 
Fig. 4. MBB in KG226-70 H. maculata. Moddergat. 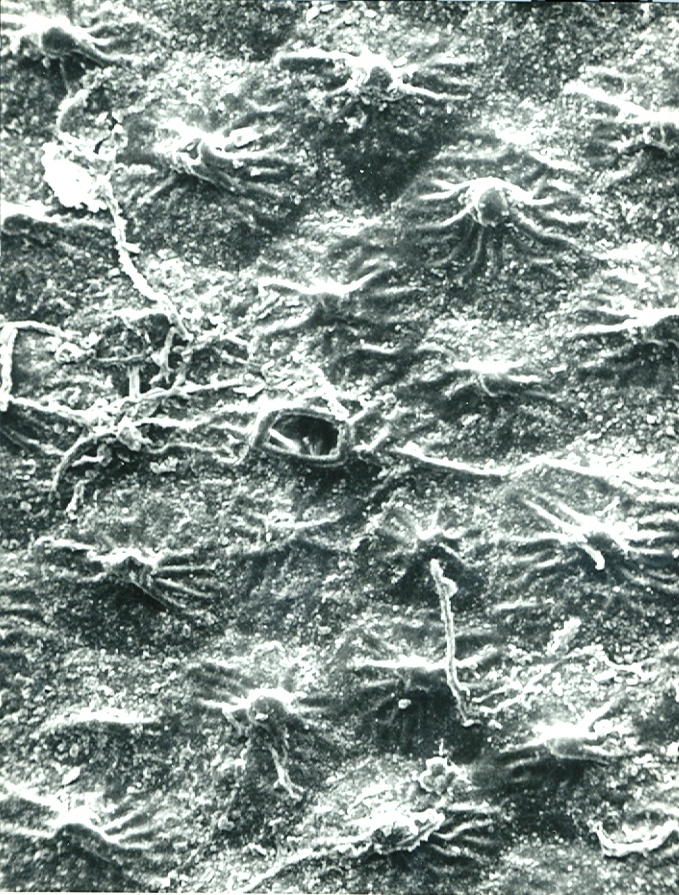
Fig. 5. MBB in KG226-70 H. maculata. Moddergat. 
Fig. 6. MBB in KG167-70 H. maculata. Brandvlei. 
Fig. 7. MBB in KG 167-70 H.maculata. Brandvlei. 
Fig. 8. MBB in KG20-70 H. maculata. Brandvlei. 
Fig. 9. MBB in KG163-71 H. emelyae var multifolia. Springfontein. 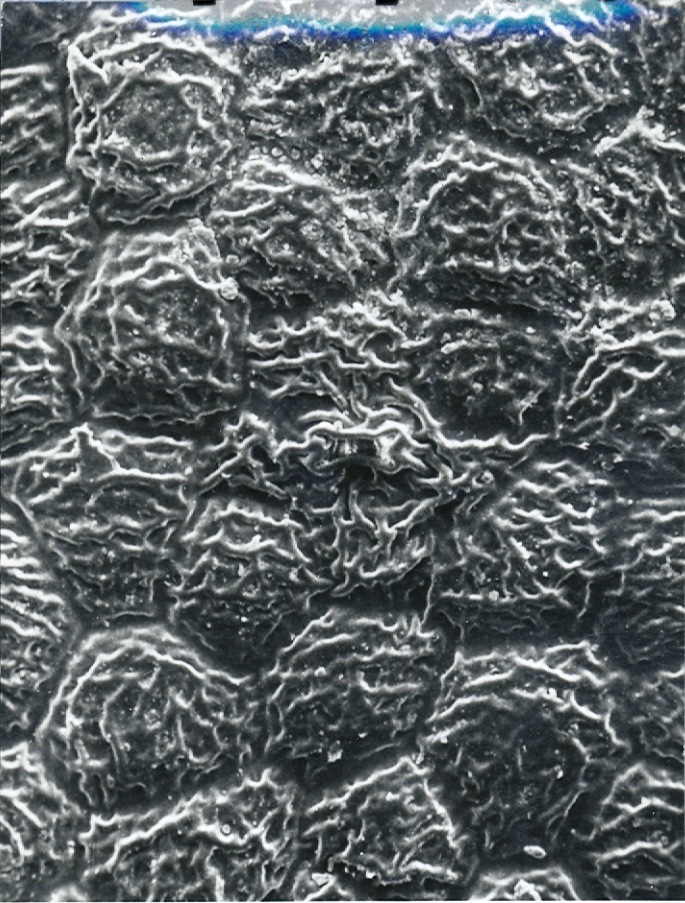
Fig. 10. MBB in KG178-70 H. reticulata. Wolfkloof, Robertson. 
Fig. 11. MBB in KG179-70 H. maraisii var notabilis. Wolfkloof, Robertson. 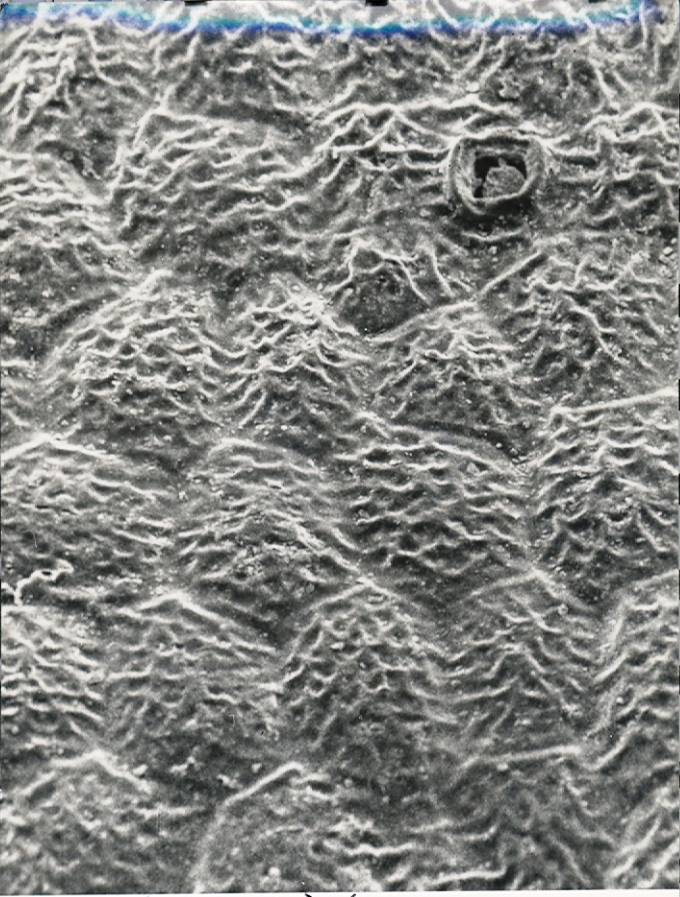
Fig. 12. MBB in KG36-70 H. heidelbergensis var minor. Rooivlei, Bredasdorp. 
Fig. 13. MBB in KG36-70 H. heidelbergensis, Rooivlei, north of Bredasdorp. 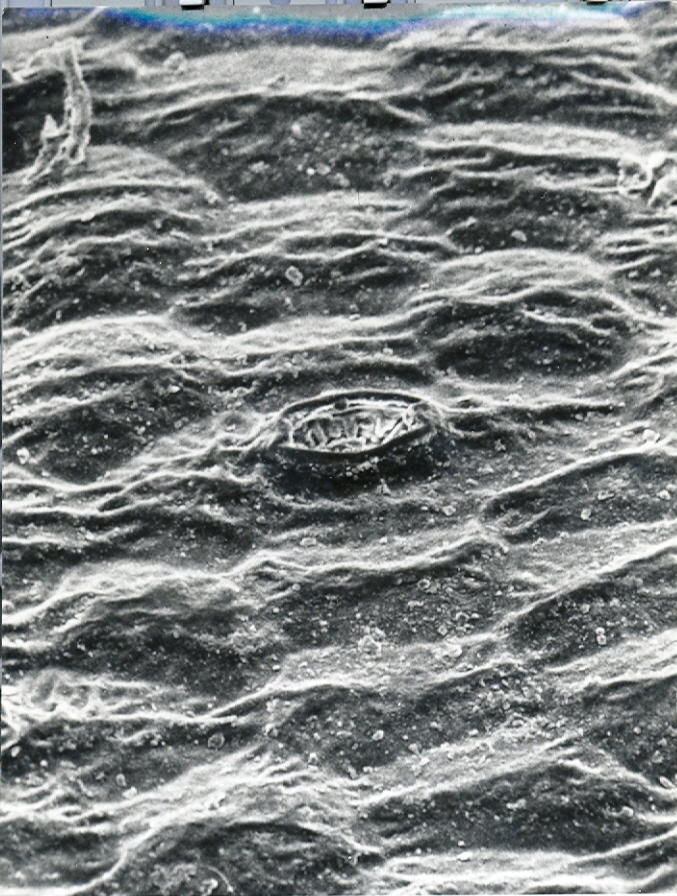
Fig. 14. MBB in KG318-71 H. rossouwii var rossouwii. Oudekraal. 
Fig. 15. MBB in KG709-60 H.mirabilis var sublineata. S Bredasdorp. 
Fig. 16. MBB in KG311-71 H. mirabilis va sublineata. N. Bredasdorp. 
Fig. 17. MBB in KG81-71 H. heidelbergensis var toonensis. Matjestoon. 
Fig. 18. MBB in KG210-71 H. heidelbergensis vr heidelbergensis. E Heidelberg. 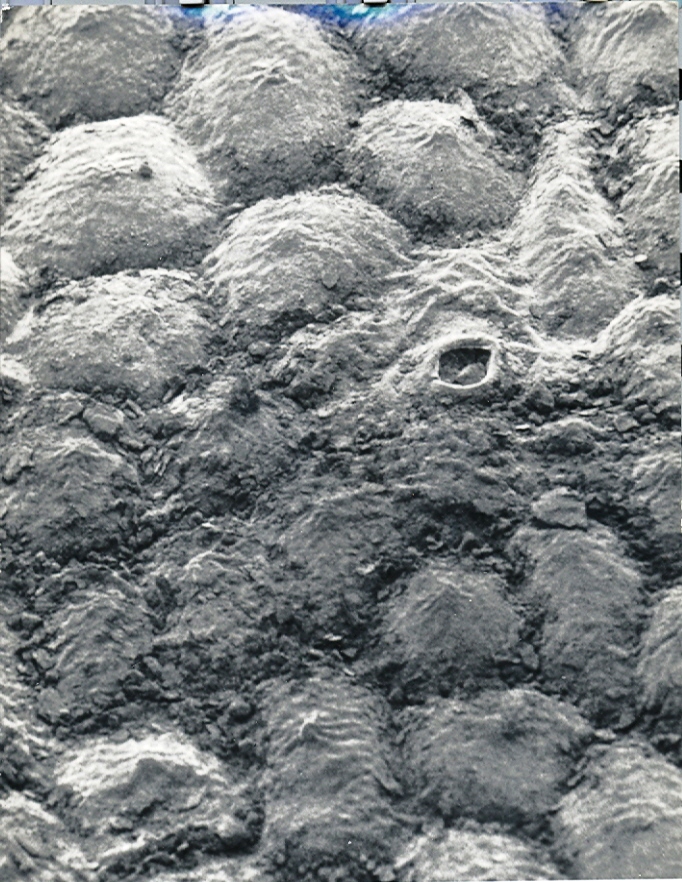
Fig. 19. MBB in KG134-70 H. mirabilis var triebneriana. Olifantsdoorn, SE Macgregor. 
Fig. 20. MBB in KG213-70 H. maraisii var maraisii. S Macgregor. 
Fig. 21. MBB in KG35-70 H. maraisii var maraisii. Rooivlei, N Bredasdorp. 
Fig. 22. MBB in KG 287-72 H. arachnoidea va nigricans. S Calitzdorp. 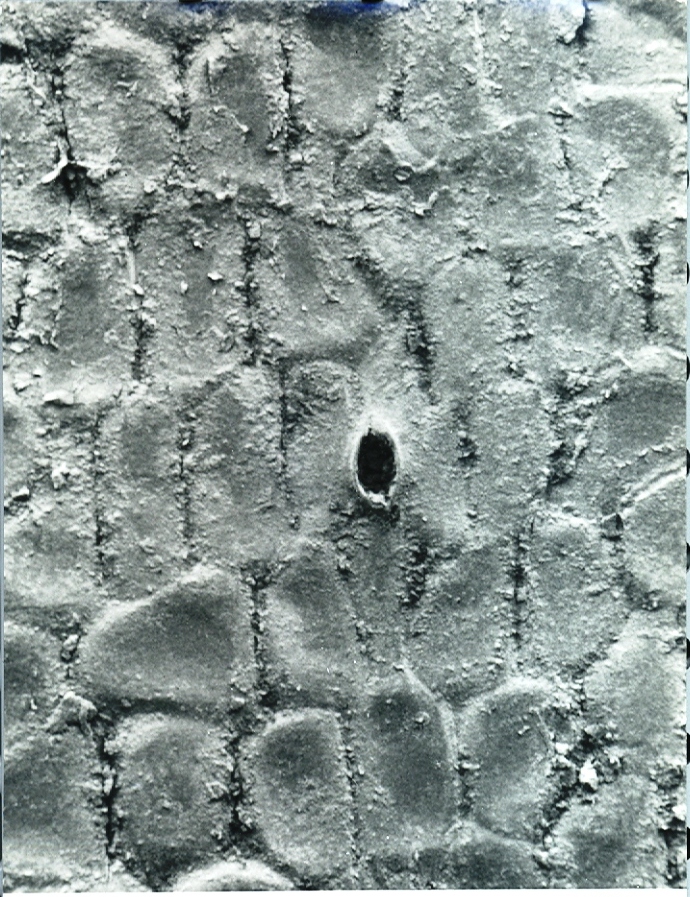
Fig. 23. MBB in KG400-61 H. maraisii var maraisii. Dublin, Worcester. 
Fig. 24. MBB in KG179-70 H. maraisii var notabilis. Wolfkloof, Robertson.
Figure captions:-
Fig.1 KG226/70 H. maculata, Moddergat, south of Worcester.
Fig.2 KG48/70 H. maraisii, Stormvlei, south of Bonnievale.
Fig.3 KG20/70 H. maculata, Brandvlei Dam, south of Worcester
Fig.4 KG226/70 H. maculata, Moddergat, south of Worcester.
Fig.5 KG226/70 H. maculata, Moddergat, south of Worcester.
Fig.6 KG167/70 H. maculata, Brandvlei Dam, south of Worcester.
Fig.7 KG167/70 H. maculata, Brandvlei Dam, south of Worcester.
Fig.8 KG20/70 H. maculata, Brandvlei Dam, south of Worcester.
Fig.9 KG163/71 H. emelyae var. multifolia, Springfontein, Little Karoo.
Fig.10 KG178/70 H. reticulata, Wolfkloof, north of Robertson.
Fig.11 KG179/70 H. notabilis, Wolfkloof, north of Robertson.
Fig.12 KG36/71 H. heidelbegensis, Rooivlei, north of Bredasdorp.
Fig.13 KG36/71 H. heidelbegensis, Rooivlei, north of Bredasdorp.
Fig.14 KG318/71 H. rossouwii, Oudekraal, south-west of Heidelberg.
Fig.15 KG709/60 H. mirabilis var. sublineata, south of Bredasdorp.
Fig.16 KG311/71 H. mirabilis var. sublineata, north of Bredasdorp.
Fig.17 KG81/71 H. heidelbergensis, Matjestoon, south of Heidelberg.
Fig.18 KG210/71 H. heidelbergensis, east of Heidelberg.
Fig.19 KG134/70 H. mirabilis var. triebneriana (the forma diversifolia) from east of McGregor.
Fig.20 KG213/70 H. maraisii (var. whitesloaneana), south of McGregor.
Fig.21 KG35/70 H. maraisii, north of Bredasdorp.
Fig.22 KG287/72 H. arachnoidea var. nigricans from south of Calitzdorp.
Fig.23 KG400/61 H. maraisii, Dublin west of Robertson.
Fig.24 KG179/70 H. notabilis, Wolfkloof, Robertson.
Appendix:-
List of specimens submitted and scanned (bracketed numbers are the laboratory codes):-
520/60 (40) H. maraisii, Cogmanskloof.
709/60 (54) H. mirabilis var. sublineata, S. Bredasdorp.
400/61 (31) H. maraisii, Dublin.
624/69 (51) H. mirabilis var. mirabilis, Mierkraal.
625/69 (58) H. mirabilis var. badia, Napier.
629/69 (47) H. maraisii, W. Swellendam.
638/69 (39) H. maraisii var. notabilis, Klaasvoogds.
661/69 (61) H. maraisii var. notabilis, Wolfkloof, Robertson.
679/69 (62) H. mirabilis var. badia, Napier.
681/69 (59) H. mirabilis, N. Napier.
688/69 (32) H. maraisii, Oliva, Robertson.
689/69 (30) H. maraisii, SW Robertson.
692/69 (53) H. mirabilis var. triebneriana, W. Genadendal.
20/70 (26) H. maculata, Brandvlei West.
26/70 (55) H. mirabilis, Stormsvlei Pass.
2?/70 (64) H. mirabilis, S Greyton.
30/70 (60) H. mirabilis, SE. Greyton.
31/70 (63) H. mirabilis, S. Greyton.
32/70 (57) H. mirabilis var. beukmannii, Skuitsberg.
35/70 (20) H. maraisii, Rooivlei, Bredasdorp.
36/70 (6) H. heidelbergensis var. minor, Rooivlei, N.Bredasdorp.
48/70 (18) H. maraisii, Stormsvlei.
106/70 (45) H. maraisii var. notabilis, Buitenstekloof.
112/70 (13) H. pubescens, Sandberg, Worcester.
134/70 (28) H. mirabilis, Olifantsdoornkloof.
167/70 (24) H. maculata, Brandvlei.
178/70 (42) H. reticulata, Wolfkloof, Robertson.
179/70 (43) H. maraisii var. notabilis, Wolfkloof.
180/70 (44) H. maraisii var. notabilis, Wolfkloof.
202/70 (7) H. magnifica var. atrofusca, Spitzkop, Heidelberg.
212/70 (27) H. maraisii, SW McGregor.
213/70 (29) H. maraisii, Houtbaaikloof.
213/70 (29b)H. maraisii, S McGregor.
223/70 (33) H. maraisii var. meiringii, W Bonnievale.
224/70 (34) H. maraisii var. meiringii, E Bonnievale.
226/70 (25) H. maculata, Moddergat.
402/70 (2) H. springbokvlakensis, Springbokvlakte.
2/71 (35) H. maraisii var. meiringii, W Bonnievale.
9/71 (37) H. maraisii var. meiringii E Bonnievale.
13/71 (38) H. maraisii var. meiringii, Goudmyn.
80/71 (11) H. maraisii, Napky.
81/71 (4) H. heidelbergensis var. toonensis, Matjestoon.
83/71 (16) H. magnifica, Riversdale Nature reserve.
84/71 (17) H. mirabilis var. paradoxa, Vermaaklikheid.
118/71 (19) H. emelyae var. major, Muiskraal.
163/71 (1) H. emelyae var. multifolia, Springfontein.
170/71 (41) H.maraisii, N. Ashton.
210/71 (5) H. heidelbergensis, NE Heidelberg.
310/71 (52) H. mirabilis var. triebneriana, Skietpad.
311/71 (56) H. mirabilis var. sublineata, N. Bredasdorp.
318/71 (3) H. rossouwii, Oudekraalkop.
326/71 (12) H. mirabilis/maraisii, Rondeheuwel.
335/71 (8) H. emelyae, Rooiberg.
344/71 (46) H. maraisii, Langvlei.
481/71 (10) H. mirabilis var. triebneriana, Leeurivier (ex van Breda).
94/72 (21) H. heidelbergensis, Leeurivier.
111/72 (14) H. bayeri, S Oudsthoorn.
114/72 (15) H. emelyae var. comptoniana, Georgida.
125/72 (49) H. arachnoidea var. nigricans, Gamka Rd.
125/72 (50) H. arachnoidea var. nigricans, Gamka Rd.
138/72 (36) H. emelyae, Sandkraal.
287/72 (48) H. mucronata, W. Ladismith.
107/74 (22) H. magnifica, Tradouw Pass.
MBB1128 (23) H. pubescens var. livida, Lemoenpoort.
‑ ‑ (9) H. mirabilis var. mirabilis, Mierkraal.