
6. Haworthia bolusii Baker, JLinn.Soc. 18:215(1880). Bayer :104(1976). Bayer :31(1982). Scott :71(1985). Type: CAPE-3224 (Graaff-Reinet): Graaff-Reinet, Bolus 158 (K).
bolusii: after H. Bolus
Rosette from 40-150mm φ, slowly proliferating. Leaves oblong-lanceolate, incurved, not acuminate or truncating, translucent bluish green, with spines on margins and keel >2mm long. Inflorescence robust, simple. Flowers broad and flat across upper basal part.
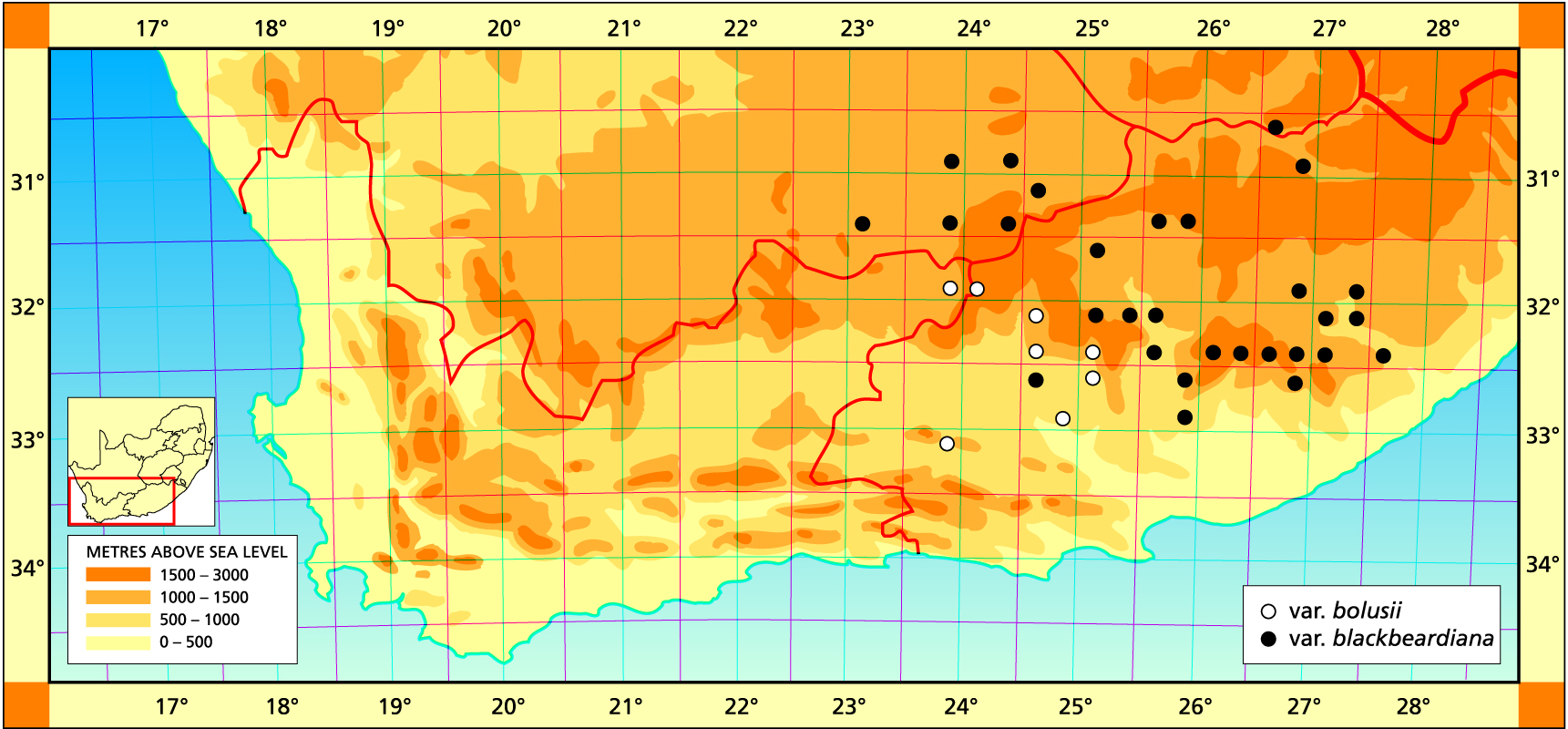
1982- The previous concept of H. bolusii was based on plants occurring around Graaff-Reinet. Here the plants are 50-60 mm in diameter and the white bristly leaves are tightly closed over the centre of the plant. The essential difference between this species and H. arachnoidea is the colour. H. bolusii has translucent bluish-green or greyish-green leaves. Further investigation in the field has shown that it is not possible to clearly separate H. bolusii and the previously recognised and larger H. blackbeardiana. Thus H. bolusii is now here considered to be a widely distributed species with a fair degree of variability. It extends from Middelburg to Sterkstroom and southwards to Graaff Reinet and Cradock. The distribution extends further southwestwards in the eastern Karoo to Klipplaat and Jansenville, where very small forms occur in some habitats. It is probable that the small, relatively hairless plants south of Klipplaat are extreme forms of H. bolusii. Similarly there are aberrant, small forms with rather fewer but larger leaves than normal in the extreme northwest near Barclay East. At Middelburg the plants are also very large (up to 150 mm diameter) as they can also be at Cradock. There is no justification for upholding H. batteniae in which the supposed distinguishing characters, viz. spiralled florets and raised leaf venation, are wholly fortuitous. The problem really lies in separating H. cooperi and H. bolusii as discussed under the former species. H. bolusii is technically in the summer rainfall region, but responds to winter growing conditions. Good light is necessary if it is to be grown to handsome natural appearances.
1999 – Col Scott’s perceptions of var. blackbeardiana, H. mucronata, H. cooperi and H. batteniae perhaps reflect the problem in this melange of species. Scott’s H. mucronata seems to include the relatively glabrous elements of three different species. His 1983 discussion of that species includes what is quite obviously the spined H. cooperi var. pilifera. My perception is that H. cooperi is a main element and includes several of Scott’s species. It is in fact difficult to exclude H. bolusii if the var. blackbeardiana is placed with cooperi; unless a distinction is made between elements which tend to have at least partially truncated leaf ends. My dilemma is that blackbeardiana could have been placed as a variety of H. cooperi as there is unquestionably a close alliance. However, this is largely a problem of the nomenclatural system. Had H. cooperi not been described, it would have not been presented here as a species but rather as a variety of a more substantial species. I concluded that larger elements with the more lanceolate leaves and longer spines from the higher altitudes of the Eastern Cape formed a more logical and coherent entity. In the northern Transkei (higher altitude) there appear to be elements with truncated leaves which I would assign to H. cooperi var. obtusata).
A similar problem arises in the wider Port Elizabeth area where a similar transition to H. decipiens occurs. I have experienced difficulty with plants collected east of Pearston but have never had adequate material on which to pass an opinion.
a. var. bolusii.
This typical variety is a smallish element in which the spination is dominant. The spines are fine and fully cover the plant so that it appears like a white ball. There is no clear integration with H. semiviva, but near Murraysburg and also at Victoria West the plants do show some tendency for leaf tips to die back as in that species. The collection from Jansenville cited below has understandably been confused with H. arachnoidea var. aranea because of the fine spination. It does have the bluish coloration of bolusii. A short distance away, on dolerite soils as opposed to tillite, the plants are larger and more typical of this species too.
Distribution:
3123 (Richmond): E. Murraysberg (-DD), Bayer 2389; Murraysburg (-DD), Banks (BOL). 3124 (Hanover): W. Graaff- Reinet (-CC), Bayer 2380 (NBG). 3224 (Graaff-Reinet): W. Graaff-Reinet (-BA), Smuts (NBG); Graaff-Reinet (-BA), Bolus 158 (BOL, K), Stayner in KG 232/62, (NBG), Ryder in NBG3152/34 (NBG); Bayer 2022, 2072 (NBG); Adendorp (-BC), Stayner in KG284/62 (NBG); Lootskloof (-DD), Bayer 2071 (NBG). 3225 (Somerset East): NE. Ashbourne (-AC), Bayer & Bruyns 6566 (NBG); NE. Pearston (-CA); Bayer 173 (NBG). 3323 (Willowmore): NE. Fullarton (-BB), Bayer 4160 (NBG); Eensaam (-BB), Marx 56 (NBG).

Haworthia bolusii var. bolusii JDV97/71 south of Graaff-Reinet. A small compact plant with fine spination. 
Haworthia bolusii var. bolusii JDV87/186 south-west of Graff-Reinet. As one moves further south and eastwards, the plants seem to lose their compactness and dense spination.
b. var. blackbeardiana (V.Poelln.) Bayer
Bayer :103(1976). Bayer :31(1982): H. blackbeardiana V.Poelln., Feddes Repert.Spec.Nov. 31:82(1932). Type: Cape, Bowes Farm near Queenstown, Miss G. Blackbeard. Not preserved. Lectotype (B&M): ex cult. V.Poelln. 1932 (B): H. blackbeardiana var. major V.Poelln. 41:196(1937). Type: Cape, Halesowen, Cradock, R. James in Parks 1483/36. Not preserved. Neotype (designated here): CAPE-3225 (Somerset East): beyond Halesowen station (-BA), Smith 2301 (NBG): H. inermis idem. 31:85(1932): H. altilinea var. inermis idem. 41:194(1937). H. altilinea var. limpida fa inermis idem. 49:29(1940). Type: Cape, Halesowen, Cradock (-BA), Stell. Not preserved. Neotype (designated here): CAPE-3225(Cradock): Halesowen (-BA), Smith 2289 (NBG): H. batteniae Scott, Cact.Succ.J(U.S.). 51:268(1979). Type: CAPE-3225(Cradock):, 35km west of Cradock (-AA), C.L. Scott 5272 (PRE): H. cooperi Bak. pp. Scott ibid 53:70(1981) as to Cradock and Thomas River. Scott :104(1985): H. mucronata Haw. pp. Scott ibid. :80(1986) as to Cradock specimen.
blackbeardiana: in honour of Miss G. Blackbeard.
This variety is generally larger than the typical form and the spination is sparser. However it is more densely spined than is H. cooperi, and the spines are longer, exceeding 2mm. The definition is otherwise geographic and the interface with H. cooperi is from south of Cathcart in a southwesterly direction to at least Jansenville. In the general Rippon Station area there are two adjacent populations, one of which is attributable to this species and the other to H. cooperi. The Patryshoogte/Baviaanskranz collection is also this species growing in the same general area of H. cooperi var. dielsiana. The interpretation in this work of H. pringlei Scott as a variety of H. decipiens may be erroneous as it may perhaps be better placed with var. blackbeardiana. This is also suggested by the Patryshoogte specimens.
There are forms of var. blackbeardiana from the Queenstown area which have very few or sparse short spines, and similarly on the Hogsback mountain. It has been reported that var. blackbeardiana has been collected in the southern Orange Free State and that its distribution extends considerably westward from Middelburg. I have no confirmation of this. There are also reports that it intergrades with H. semiviva in the far west of its range, and the sparse herbarium record suggests that this may be true.
Distribution:
3023 (Britstown): Die Puts (DD), Fuller 193 (BOL). 3024 (Philipstown): Hanover to De Aar (-CD), Stayner in KG307/70 (NBG); 3026 (Aliwal North): Elandshoek (-DA), Smith 6828 (NBG); SE. Aliwal North (-DD), Smith 7471 (NBG). 3123 (Richmond): Richmond (-AC), Stayner in KG591/69 (NBG); Richmond (-BD), Wohlman in KG581/69 (NBG). 3124 (Hanover): Cypherwater (-AD), Bruyns 3010 (NBG); Bethesda Road to Goliadskraal (-BA), Smith 3640 (NBG). 3125(Steynsberg): Thebus (-BC), Smith 3635 (NBG); 8km E. Steynberg (-BD), Smith 3634 (NBG); Tafelberg (-CA), Smith 3828 (NBG), Stayner in KG693/71 (NBG). 3126 (Queenstown): Bowes’ farm (-DD), Britten 160 (BOL), Bayer in KG 312/70 (NBG); Finchams Nek (-DD), Smith 7212 (NBG); Nonesses Nek (-DD), Smith 7216, 7217 (NBG); Longhill (-DD), Bursey in KG393/70 (NBG). 3127 (Molteno): Cofimvaba (-CD), Bruyns 4385 (BOL). 3224 (Graaff Reinet): Kendrew (-DA), Rossouw 183 (NBG). 3225 (Somerset East): 35km west of Cradock (-AA), C.L. Scott 5272 (PRE); Doornberg (-AA), Herre STE6632 (BOL); Denmark (-AB), Bruyns 1769 (NBG); Post Chalmers (-AB), Smith 7444 (NBG); 24km SE. Fish River Stn. (-BA), Smith 5791 (NBG); beyond Halesowen station (-BA), Smith 2301 (NBG); Halesowen (-BA), Smith 2289, 2301, 5340 (NBG), Herre STE6668 (BOL); Cradock (-BA), Smith 2263, 2289 (NBG), Britten (BOL); Swagershoek Pass (-BC), James 474 (BOL), Fourcade 40 (NBG), Bruyns 1627 (NBG); Daggaboer (-BC), James (BOL); Baviaanskranz, Patryshoogte (-DB), Bayer & Bruyns 6561 (NBG); NW. Rippon Stn. (-DD), Bayer & Bruyns 6556 (NBG). 3226 (Fort Beaufort): Huntley Glen (-AC), Smith 7493 (NBG); Austrey (-AD), Branch 370 (NBG); Katberg (-BC), Luckhoff in NBG407/34 (NBG); Waterdown Dam (-BD), Bayer & Bruyns 6569 (NBG); S. Estrelle (-BD), Bayer & Bruyns 6570 (NBG); Hogsback (-DB), Smith 375 (NBG). 3227 (Kingwilliamstown): Imvani (-AA), Bayer in KG311/70 (NBG); 24km N. Cathcart (-AA), Smith 3633 (NBG); N. Waku (-AA), Smith 5718 (NBG); 2km N Goshen (-AA), Branch 20 (NBG); Turnstream (-AB), Bayer & Bruyns 6571 (NBG); New Haven (-AB), Branch 18 (NBG); W. Cathcart (-AC), Smith 5744 (NBG); 16km W. Cathcart (-AC), Smith 38, 358 (NBG); SW. Cathcart (-AC), Barker 3427 (NBG), Bayer in KG310/70, KG393/70 (NBG); Inverbolo (-BC), Bruyns (NBG).
Inadequately located: Winterberg, Tarkastad, Armstrong (BOL); Queenstown, Ingram 1509 (BOL), Galpin 263 (BOL), Dyer 8 (BOL).

Haworthia bolusii var. blackbeardiana JDV91/102 soth of Cradock. This variety is widespread and the spination is coarser and sparser, or occasionally absent. 
Haworthia bolusii var. blackbeardiana JDV91/99 south of Cathcart. Here the plants are often sparsely spined and easily confounded with H. cooperi. 
Haworthia bolusii var. blackbeardiana JDV97/62 Aliwal North. The variety is not well-known at its north-eastern limits.