Printed in National Cactus and Succulent Journal, 27:51(1972).
Part 2. HAWORTHIA HERBACEA (Miller) Stearn.
M. B. Bayer, National Botanic Gardens of South Africa, Karoo Gardens, Worcester.
Abstract: The application of the name Haworthia herbacea (Mill.) Stearn to field populations is discussed. It is concluded that this name applies best to the predominant complex occurring in the western and south-western Robertson Karoo. The species H. aegrota v Poelln., H. submaculata v Poelln., H. luteorosa Uitew. and the variety H. pallida var. paynei are shown to have their origin in the same complex, and are therefore reduced to synonymy.
15. Haworthia herbacea (Mill.) Stearn, Cactus J. 7:40(1938). Bayer, Nat.Cact.Succ.J 27:51(1972). Bayer :122(1976). Bayer :42(1982). Aloe herbacea Mill., Gardeners Dictionary :n18(1768). Type: icon, 130:t131 Boerhaave Index Alter Pl. (1720). Epitype (B&M): CAPE-3319(Worcester): N. Ribbokkop (-DC), Bayer 161 (NBG): A. atrovirens D.C., Pl.Gras. :f81(1799). H. atrovirens (D.C.) Haw., Revis. :57(1821). Type: as above: A. arachnoidea var. pumila Aiton, Hort.Kewensis 1:468(1789). Willd., Spec.Pl. 2:188(1799). A. pumila (Willd.) Haw., Trans.Linn.Soc. 7:10(1804). H. pumila (Willd.) Duval, Pl.Succ.Hort.Alenc. :7(1809). Haworth, Syn.Pl.Succ. :95(1811). Type: as above (Linnaeus, Aloe pumila var. E): Aloe translucens Haw., Trans.Linn.Soc. 7:10(1804). H. pellucens Haw., Syn.Pl.Succ. :96(1812). Aloe arachnoidea var. translucens (Haw.) Ker-G., Curtis’ Bot.Mag. t.1417(1811). H. translucens Haw., Suppl.Pl.Succ. :52(1819). H. arachnoidea (L.) Duv. sensu Scott, Cact.Succ.J(U.S.) 49:205(1977). Scott, Aloe 16:41(1978). Scott :39(1985). Type: Cape, Masson. Not preserved. Lectotype (designated here): icon. :t.1417 Curtis’ Bot. Mag.: H. pallida Haw., Revis.:56(1821). Type: Not preserved. Lectotype (designated here): icon (K): H. aegrota V.Poelln., Desert Pl.Life 11:193(1939). Type: Cape, Worcester, Swellendam etc. H. Venter. Not preserved. Lectotype (designated here): icon (B): H. submaculata idem. Type: Cape, Worcester, etc. H. Venter 5. Not preserved. Lectotype (designated here): icon (B): H. luteorosea Uitew., Cact.en Vetpl. 5:88(1939). Type: ex Hort, Holland. Not preserved.H. arachnoidea (L.) Duval sensu Scott, Cact. Succ. J. (US) 49:205 (1977).
herbacea: yellow-green.
Rosette stemless, proliferating, to 8cm φ. Leaves erect, incurved, scabrid, margins and keel with firm spines, greenish yellow in colour, reticulate patterning with translucent interstices. Inflorescence simple, to 300mm. Flowers large, beige, pinkish tips, buds bi-arcuate.
1982 – The choice of the name herbacea is possibly a poor one as this name is associated with a number of perhaps incompatible early illustrations. Scott’s selection of the name arachnoidea is equally dubious particularly as it also ignores the flower character. H. herbacea occurs only in the Worcester area extending in the southeast to east of McGregor, south into the mountains at Villiersdorp where a large flowered form occurs and to just northwest of Worcester itself. It is very close to H. reticulata and southwest of Robertson it is often difficult to know with which species one is dealing. However, H. herbacea usually has beige flowers, the plants are usually solitary, more deeply sunk in the ground, less proliferous and more hairy than H. reticulata. At De Wet the two species grow together and there are distinct hybrids. There are at least two other localities where both species grow in close association without hybridisation. H. herbacea flowers in late winter (September‑October) and has an unusually large flower. The Villiersdorp mountain form collected by E. Esterhuysen, has a particularly impressive flower but has not been introduced into cultivation. H. herbacea has a direct connection with H. maculata at the Brandvlei Dam south of Worcester. The size of the plant varies from 80mm in diameter to as little as 35mm near McGregor where the flowers may also be pinkish.
1999 – Nomenclature is indeed a slippery slope and this is a case in point. The name pumila which Duval applied in Haworthia, was based on Boerhaave’s early illustration as can be traced through a tortuous synonymy. It is in my opinion now invalid in the genus Haworthia, because the same type is used for the name herbacea. (see also Haworthia margaritifera). Apart from this nomenclatural tangle, the application of the name to a naturally occurring element is a problem. The Boerhaave illustration can perhaps be better related to Haworthia maraisii than to the species for which I have used the name. The earlier epithet atrovirens would have been a far better choice for that species had it been available. Rather than make this unfortunate change, and then also using the name translucens for this species, I am following a principle of conservation and letting sleeping dogs lie. It is enough that Col Scott has unfortunately confounded the issue by his use of the name H. arachnoidea for this species, without the acknowledgment in synonymy that may have clarified the issue.
The type nominated by Breuer and Metzing for H. translucens is based on the 1982 Handbook interpretation, where the name was applied incorrectly. It is apparent that H. translucens Haw. as perceived in 1819, includes the Botanical Magazine illustration cited above. This must be related to H. herbacea as understood here now.
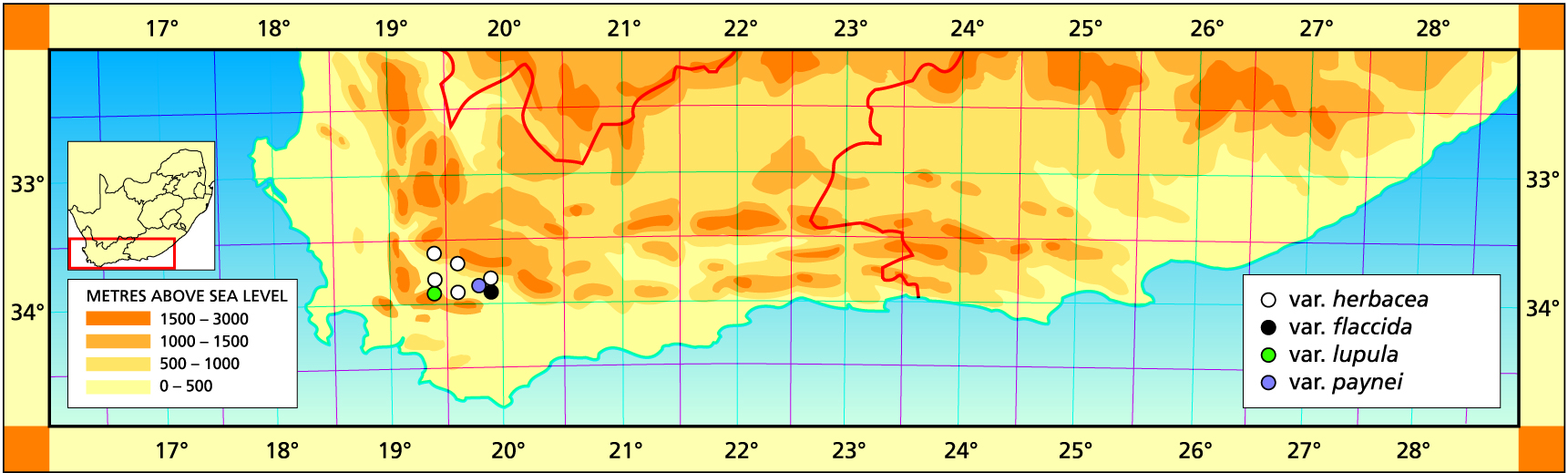
a. var. herbacea. The typical variety is widespread and relatively homogeneous throughout its range.
Distribution: 3319 (Worcester): Karoo Garden (-CB), Dobay 62 (NBG), Barker 8766 (NBG); Veld Reserve (-CB)., Olivier 129 (NBG, PRE), Pamphlet 44 (NBG); W. Worcester (-CB), Searle in NBG162/84 (NBG), Bayer 2421 (NBG); Brandwacht (-CB), Bayer in KG166/70 (NBG); SE. Brandvlei Dam (-CB), Bayer 2422 (NBG); W. Doornrivier (-CD), Bayer 1995 (NBG), Leipoldt (BOL); Lemoenpoort (-CD), Bayer 1996(NBG); N. Lemoenpoort (-CD), Bayer 4439 (NBG); Patryskloof, Mauve & Oliver 243 (NBG); De Wet (-DA), van Heerden 1024 (NBG); Rabiesberg (-DA), Esterhuysen & Lewis in NBG2689/35 (NBG); Ribbokkop (-DC), Bayer 161 (NBG); Keerweerder, Jonaskop (-DC), Bayer 2697 (NBG); Nuy (-DC), Hurling & Neil (BOL); Mowers (-DC), Bayer in KG218/70 (NBG); Wansbek (-DC), Bayer 1997 (NBG), Moffett in KG181/70; Rabiesdal (-DD), Heunis 1 (NBG); Koningsrivier (-DD), Bayer in KG 329/70 (NBG).
Inadequately located: Marloth 8912 (PRE); Worcester, Smith 3910 (NBG), Hurling & Neil (BOL), Venter 4 (BOL), Joubert 1 (BOL); Worcester to Robertson, Venter 5, 7 (BOL); ex hort, Leipoldt (BOL), Logan (BOL), Starke in BOL24591, Venter in NBG215/39, Aryer’s nursery (BOL); Ross-Frames in NBG78/44; Bonnievale, Malherbe in NBG297/40, van der Merwe 174 (BOL); Riversdale, Ferguson (BOL).
Haworthia herbacea var. herbacea JDV84/5 Ribbokkop. Grows, and hybridises with H. reticulata at this site.
Haworthia herbacea var. herbacea JDV85/60 Bosfontein. The spination of the plants in this area is more pronounced.
Haworthia herbacea var. herbacea JDV85/12 Sandberg. Here it is growing in quartzitic rock.
b. var. flaccida var.nov. Type: CAPE-3319 (Worcester): Rooiberg (-DD), P.V.Bruyns 7114 (NBG, Holo.).
flaccidus: weak, soft.
Differs from the species in being small and delicate. There is considerable variation among the plants in habitat. (A var. herbacea foliis parvioribus et virellis differt).
The habitat of this variety is on steep north-facing rocks where the plants occur in small very dense clusters. They co-occur with H. maraisii and with H. reticulata. The former is on the same rock face but not as exposed, while the latter occurs on slopes both lower down and higher up on the mountainh. The locality is outside the known northeast distribution of the species. There are very few plants and the habitat is uncharacteristic of any of the species in the Southern Cape. The flower is outwardly identical to that of H. herbacea as is also that of H. pubescens. The flowering time co-incides with that of the latter but there is very little vegetative similarity.
Haworthia herbacea var. flaccida JDV96/31 Rooiberg. Still very poorly known from only four clones.
Haworthia herbacea var. flaccida JDV96/31 Rooiberg. This clone more closely resembles the typical variety.
c. var. lupula var.nov. Type: CAPE-3319 (Worcester): Boscheveld Mt., Wolfkloof, Villiersdorp (-CD), E. Esterhuysen (NBG, Holo.).
lupula: small wolf as reference to origin.
Distinctive for the larger pink flower, and for the broader, shorter leaves which are more finely flecked and slightly less scabrid than the typical.
This variety was first collected by Elsie Esterhuysen who specialised in the flora of the Cape mountains. It occurs in sandstones, which is not unique for this species as it is also in sandstones south of McGregor. The occurrence is at the southwestern limits of the species. This is one of the elements which recurs through the Cape mountains and which suggest a common ancestry from here for at least the subgenus Haworthia. The similarities to H. vlokii, H. mirabilis var. consanguinea, H. maculata, H. variegata var. modesta (on the Potberg Mountain) and to H. turgida at high altitude are notable.
Haworthia herbacea var. lupula JDV94/36 Villersdorp. A surprising occurrence in sandstone with Fynbos vegetation.
Haworthia herbacea var. lupula JDV94/36 Villiersdorp. The flowers of this variety are surprisingly large and pinkish.
d. var. paynei (V.Poelln.) Bayer comb.nov. H. paynei V.Poelln., Feddes Repert.Spec.Nov. 41:206(1937). H. pallida var. paynei V.Poelln., Cactus J 6:19(1937). Type: Cape, McGregor district, G. Payne. Not preserved. Neotype (designated here): CAPE-3319 (Worcester): Olifantsdoorn, McGregor (-DD), Bayer 4430 (NBG).
paynei: for G. Payne.
Mr George Payne accumulated an extraordinary collection of plants prior to the war and subsequently lost interest. This variety named after him is indeed different for its small size and for the bi-coloured flower which is pink above and white below. It is on the southeastern limits of the species and a still smaller proliferous form has been reported in the area.
Distribution: 3319 (Worcester): McGregor (-DD), Payne in PRE 27254, in PRE 34948, Bayer 695/69 (NBG); 1km S. McGregor (-DD), Bayer (NBG); Near McGregor (-DD), Smith 3978, 7196 (NBG); 3km E. McGregor (-DD), Scott 2203 (PRE); Olifantsdoorn, McGregor (-DD) Bayer 4430 (NBG).
Haworthia herbacea var. paynei JDV96/41 McGregor. A smaller plant and the flowers have pinkish upper lobes with traces of green, whereas the lower lobes are whiter
Set 10. Map point 18. H. herbacea ‘submaculata’. Brickfield, Brandvlei Dam. Figs. 10.1 to 10.20 MBB7995 H. herbacea, S Brandvlei Brickfield.
Figs. 10.21 to 10.53 MBB7996 H. herbacea, E Brandvlei Brickfield.
I have associated this locality with vonPoellnitz H. submaculata and treated it as a synonym of H. herbacea. However, here I first illustrate a population about 600m south of that where the plants are in the usual size range for the species ie.30-40mm diam. At the locality east of the Brickfield and next to the Breede River, the plants are 1/3 to 1/2 as large again and can form huge clumps. North-west from this is a population of H. maculata at the extreme end of Die Nekkies and only about 300m distant, and this population I have always regarded as somewhat intermediate. What is interesting to note is the huge variation in leaf shape and armature within each population. In both cases the plants are in Witteberg Sandstones and at the first site this is both in a very shale-like stratum as well as in a highly quartzitic one. This is unusual for H. herbacea. Both populations wedge in geographically between H. maculata populations and in no known case do both occur.
I need to point out that there is a still earlier article which covers Haworthia maculata (Haworthia maculata <–> Haworthia pubescens) that lays the basis for this discussion. In that article I note the position of the Sandberg to Cilmor and DeWetsberg and intended to include the Sandberg H. pubescens in that article. We could not get landowner contact and so that fell away. However, this problem was overcome and we first explored a Dwyka Tillite outcrop southeast of Sandberg. There is a vast accumulation of windblown sand on the first hill and we saw no Haworthia. There is a smaller hill further to the southeast that is also Dwyka and erosion exceeds wind deposition so smaller non-geophytes do quite well. We found both H. herbacea (see fig.1 MBB8014) and H. pumila there. From there we went to the southernmost point of the Sandberg. A misjudgement landed our vehicle in mud and the drama to get out limited the time we had to explore. We found a lone H. herbacea (fig. 2 MBB8012). Returning a week later we approached the Sandberg from the southwest, and almost immediately on reaching the top we found H. pubescent. Fig. 3 is a view towards Cilmor and DeWetsberg where the plants appear to be intermediate H. pubescens↔H. maculata. The picture is useful to get some idea of the role of geographic and geological considerations. The high mountains in the background are Table Mountain Sandstone and no Haworthia is known there. I am not certain that this is true and G J Payne did tell me that he had seen plants on the extreme lower right and south of the Brandvlei Dam. But also on the absolute distant and absolute left, is the Riviersonderend Mt. That is also TMS. The deep Wolfkloof Valley behind that is the locality for the much unexpected H. herbacea ‘lupula’. (These inverted single commas are not entirely necessary but I use them to underscore my informal use of names that have less reality. The var. lupula is real). The mountains ahead of that last line are Hammansberg on the left and the Moddergat to the right. Between there and DeWetsberg has not so far turned up Haworthia, but this is an exploration problem. Behind the DeWetsberg is also underexplored. H. herbacea does occur between DeWetsberg and behind the mountains on the low right just in the picture and also east of the brickfield out further right. H. maculata is only known in this area along the Nekkies north (further to the right) of the Brandvlei Dam just visible in the picture.
01. MBB8014 H. herbacea, E Sandberg.
02. MBB8012 H. herbnacea, Klipkranz Camp.
03 . View west of south from Sandberg.
Prior to this exploration H. pubescens was to me only known from the northern part of the Sandberg that lies south of a road going eastwards to Eilandia between Worcester and Robertson. Here H. herbacea does occur on the lower northwest warm slopes. H. pubescens seems to occur only on the upper two ridges and H. herbacea is not known to intermingle with it. This is all Witteberg Sandstone that as a formation overlies Bokkeveld Shale and underlies TMS.
Coming back to the southwest corner of the Sandberg where we found H. pubescens. The plants seem very similar to the species as it occurs to the north. They were very cryptic and often in shady rock retreats where they were really hard to see. It was mid- to late-morning when we were there and the plants were not going to be better exposed as the sun moved further west. Although there was very suitable well-drained habitat lower down on the shaded east slopes, there were no plants and I speculate that this may be because the plants may need the cooling effect of wind movement up on the ridge. The pictures tell the story of variability in respect of a whole range of leaf and rosette characters.
It is worth noting fig. 43 of the dead remains of a plant under a clump of restioid. It seems that seedling survival is closely coupled to early protection giving rise to the concept of nurse-plants. Plants are often very difficult to find because they are so hidden beneath accompanying vegetation. But they do need light and the dynamics of vegetation growth and densification must have quite a big impact on the ageing and survival of plants. It raises again the question of how long do the plants live? For plants like Aloe ferox and A. dichotoma I do have a real experience of a lower limit of about 35 years and a top limit in the several hundred. In the field, the plants seem comfortably ageless.
The really interesting part is this. While I was busy tediously cleaning a plant to photograph it, Daphne called to me to come and see a lighter green plant she had seen. Moving in that direction I saw a plant that registered as H. herbacea but with some hesitation and doubt (see fig.4). I then went to see what Daphne had observed. They seemed less obviously H. herbacea but that seemed to be a logical and conservative opinion (see figs. 5-11). That was until Daphne found two adjacent rosettes at the foot of a restioid clump that left me in no doubt that they were hybrid H. pubescens/maculate (see figs 9-12). These were in bud whereas H. pubescens plants showed no sign of impending flowering. Note the buds are less well developed than MBB8014 further east, even if possibly insignificant. Going back to the other plants we confirmed my doubts. They were a lighter colour and apparently softer texture that we would have expected in H. herbacea. These were the only plants we saw in a space bridging the occurrences of plants of H. pubescens.
04. MBB8013 H. herbaceaXherbacea, Sandberg.
05-11 MBB8013 H. herbaceaXherbacea, Sandberg
12-46 MBB8013 H. pubescens, SW Sandberg.
We did not explore the western slopes where habitat would have been more suitable for H. herbacea and I expect it does occur there. What puzzles me is that so frequently have I found very distinctive hybrids between species in close proximity and very seldom where the species are some distance from one another. I cannot say I have ever found a hybrid in the clear absence of both parents. The example of Astroworthia bicarinata at Lemoenkloof, east of Barrydale, may be an exception where only Astroloba corrugata (syn A. muricata, A. aspera) is present but H. pumila apparently not. Hybridization is thought to be an important element in the “evolution” of new species. I doubt this as it is quite evident that separation into two species is a pre-requirement. If new species have evolved in Haworthia by hybridization, how did they evolve as such in the first place? The answer to me lies in the continuities between populations. I observe, and have experienced of expected continuity between populations. While the Cilmor populations are thought to be H. pubescens↔maculata it cannot be said anymore that they are hybrid, or populations where the morph or drift to discrete elements has not reached a conclusion. The latter is more likely. As there is already apparent geographic continuity of H. maculata and H. herbacea, I was expecting some evidence of a similar relationship between H. herbacea and H. herbacea. So here it is. Hybridization as a factor in speciation in Haworthia does not seem to very likely. It confirms for me that there is a fractal “chaotic” order to species in Haworthia and the reality is that a view of many truly discrete species is a fabrication and a very ill-considered view.
Acknowledgement:
We are always greeted with such kindness and helpfulness that we might have expected this from the Sandberg landowners too. It came in no small measure. Driaan Griesel was most enthusiastic and interested and also helped us with extracting our vehicle from the mud on the one occasion, and then jump-starting it after a flat battery on the second. Our imposition did not so much as touch his view of the day.
(ed. – Bruce made another visit to Southwest Sandberg on 9 December 2012 and includes the following flower pictures. He makes this comment; personal correspondence 27 December 2012.)
I am actually not sure at all about flowering time now. I used to be quite sure of being able to collect seed of pubescens mid-Nov. But I observed at Humansdorp that gordoniana peak flowering could be out by 6 weeks. In any case the plants can produce successive spikes so one can get delayed flowering and added to that energy in the first or the second flower set. I know mirabilis at MacGregor can flower from Nov. thru to March while at Montagu mirabilis can flower as late as April/May. Retusa and geraldii are quite happy to produce flowers in either Spring or Summer and Kobus observed that splendens did that too. Maculata can flower from Sept. thru to late Dec. And each population does its own thing.