24. Haworthia mutica Haw. Revis :55(1821). Scott, Aloe 11:4(1973). Bayer :139(1976). Bayer, Excelsa 8:50(1978). Bayer :48(1982). Scott :118(1985). Type: Cape. Not preserved. Lectotype: icon (K). Epitype (B&M): Bredasdorp to Swellendam, Soesriver, Bayer in KG623/69 (NBG): H. otzenii Smith, JS.Afr. Bot. 11:72(1945). Type: CAPE‑3419 (Caledon): Bredasdorp (-BD), Otzen 6 in Smith 5478 (NBG).
mutica: without a point.
Rosette stemless. non-proliferous, 6-8cm φ. Leaves 12-15, retused, blunt-tipped, brownish-green, in habitat developing purplish cloudiness, barely pellucid with several longitudinal lines. Inflorescence simple, to 20cm. Flowers white with brownish veins.
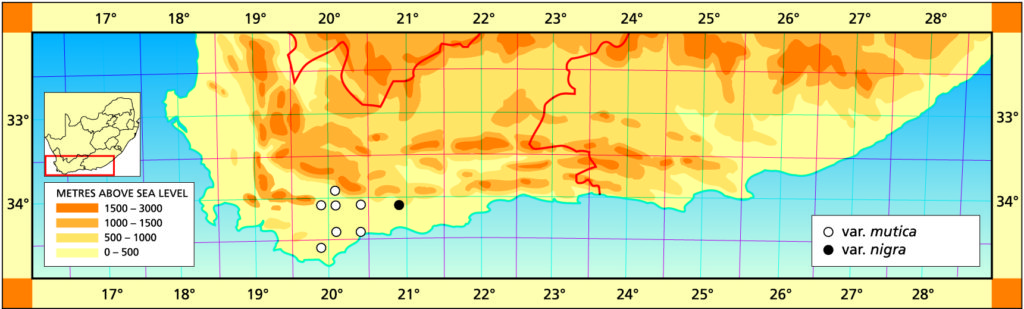
1982 – The name mutica is suggested by the leaf tips which are rounded. This species is very similar to H. retusa and if the two species do meet it can be expected to be in the area between Heidelberg and the lower Breede River. H. mutica is only known west of the Breede River south of Swellendam, but it occurs at one locality north of the river at Drew. In the field the venation and leaf colour is slightly purplish. It is interesting to note that it is very often difficult to distinguish between smooth forms of H. pygmaea and H. mutica. In one comparison of the flowers the only difference that could be detected was that the bracts were purplish‑veined in H. mutica as opposed to greenish in H. pygmaea. H. mutica leads a rather precarious existence in an intensively farmed area, and survives on rocky shale ridges. It covers nearly the same distribution range as H. mirabilis but the two species never appear to grow in the same habitats. H. mutica is generally more glaucous than H. retusa and has a characteristic bluish‑brown coloration.
1999 – A relationship has always been sought between this species and H. retusa. However, a more appropriate solution lies in the recognition that H. retusa should either be enlarged to encompass H. turgida and all its variants, or regarded as a separate species with no interaction with others in terms of its range or distribution – a difficult decision.
a. var. mutica.
This is the element west of the Breede river. It was once known from northwest of Drew (north of the Breede River) from where it was collected and sold in pre-war times. That locality has since been taken in by wheat fields, and a single solitary clone survives in the Karoo Botanic garden collection. Leaves of this clone have been propagated. It is remarkable for the white opacity which has subsequently developed in the leaves, and which is also appearing in offsets of the original clone.
Distribution: 3320 (Montagu): N. Drew (-CC), Smith 5614 (NBG); Drew (-CC), Fouche in PRE 34916. 3419 (Caledon): E. Riviersonderend (-BB), Otzen 6 in Smith 5478 (NBG); 10km E. Riviersonderend (-BB), Smith 3270, 3460, 3970, 5478 (NBG), Bayer 4479 (NBG); Klipdale (-DB), (NBG). 3420 (Bredasdorp): E. Stormsvlei (-AA), Smith 3249 (NBG); Noukloof (-AA), Bruyns (NBG); Rietfontein (-AA), Bayer 2526 (NBG); Crodini (-AB), Bayer (NBG); Soesriver (-AC), Bayer 4473, in KG623/69 (NBG); Kykoedie (-AC), Bayer in KG83/77 (NBG); N. Kykoedie (-AC), Bayer in KG327/71 (NBG); Badjieskraal (-AC), Bayer in KG85/72 (NBG); Haarwegskloof (-AD) Venter 2 (NBG).
Inadequately located: ex hort, Malherbe, Smith 3430 (NBG); Caledon, Hurling & Neil (BOL); Bredasdorp, Theunissen (BOL).

Haworthia mutica var. mutica JDV92/64 Drew. The only clone known to have survived avaricious collecting. Collected in 1970, it has developed a curious and progressive white cloudiness in the leaves – also observed, albeit rarely in other collections. 
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. This species flowers earlier than H. retusa. 
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. In the field the plants have a purplish waxy bloom to the leaves.
b. var. nigra var. nov.
Type: CAPE-3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG, Holo.).
nigra: black.
Differs from the type in its very dark coloration and its occurrence south and east of Heidelberg. (A var. mutica colore perfuscato differt).
This variety from Kransriviermond has been known for a long time and was distributed as H. retusa. It is very dark green, although this does depend on growing conditions as under softer light it may be quite green. There are also populations, probably of this variety, just east of Heidelberg.
Distribution: 3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG); Heidelberg (-BB), Smith 5509 (NBG); E. Heidelberg (-BB), Smith 5755 (NBG); 2km W. Heidelberg (-BB), Smith 6196, 6567 (NBG); N. Heidelberg (-BB), Smith 5509 (PRE).

Haworthia mutica var. nigra JDV86/16 south of Heidelberg. Less wax-like and dark green. 
Haworthia mutica var. nigra JDV90/1 east of Heidelberg. There are three such populations known which seem to link H. mutica to H. turgida and to H. retusa.