24. Haworthia mutica Haw. Revis :55(1821). Scott, Aloe 11:4(1973). Bayer :139(1976). Bayer, Excelsa 8:50(1978). Bayer :48(1982). Scott :118(1985). Type: Cape. Not preserved. Lectotype: icon (K). Epitype (B&M): Bredasdorp to Swellendam, Soesriver, Bayer in KG623/69 (NBG): H. otzenii Smith, JS.Afr. Bot. 11:72(1945). Type: CAPE‑3419 (Caledon): Bredasdorp (-BD), Otzen 6 in Smith 5478 (NBG).
mutica: without a point.
Rosette stemless. non-proliferous, 6-8cm φ. Leaves 12-15, retused, blunt-tipped, brownish-green, in habitat developing purplish cloudiness, barely pellucid with several longitudinal lines. Inflorescence simple, to 20cm. Flowers white with brownish veins.
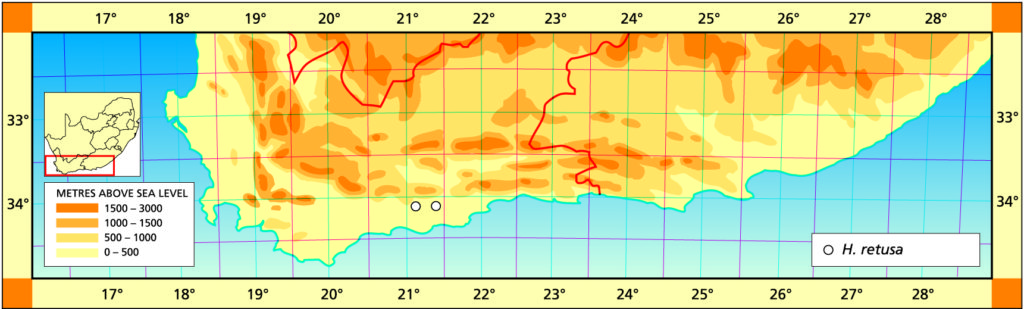
1982 – The name mutica is suggested by the leaf tips which are rounded. This species is very similar to H. retusa and if the two species do meet it can be expected to be in the area between Heidelberg and the lower Breede River. H. mutica is only known west of the Breede River south of Swellendam, but it occurs at one locality north of the river at Drew. In the field the venation and leaf colour is slightly purplish. It is interesting to note that it is very often difficult to distinguish between smooth forms of H. pygmaea and H. mutica. In one comparison of the flowers the only difference that could be detected was that the bracts were purplish‑veined in H. mutica as opposed to greenish in H. pygmaea. H. mutica leads a rather precarious existence in an intensively farmed area, and survives on rocky shale ridges. It covers nearly the same distribution range as H. mirabilis but the two species never appear to grow in the same habitats. H. mutica is generally more glaucous than H. retusa and has a characteristic bluish‑brown coloration.
1999 – A relationship has always been sought between this species and H. retusa. However, a more appropriate solution lies in the recognition that H. retusa should either be enlarged to encompass H. turgida and all its variants, or regarded as a separate species with no interaction with others in terms of its range or distribution – a difficult decision.
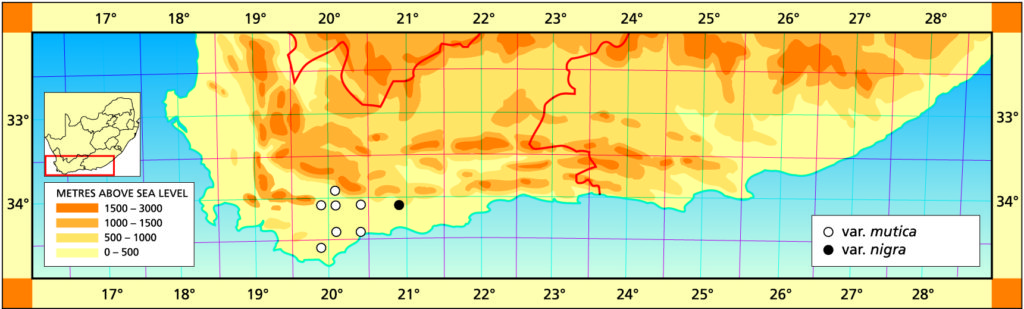
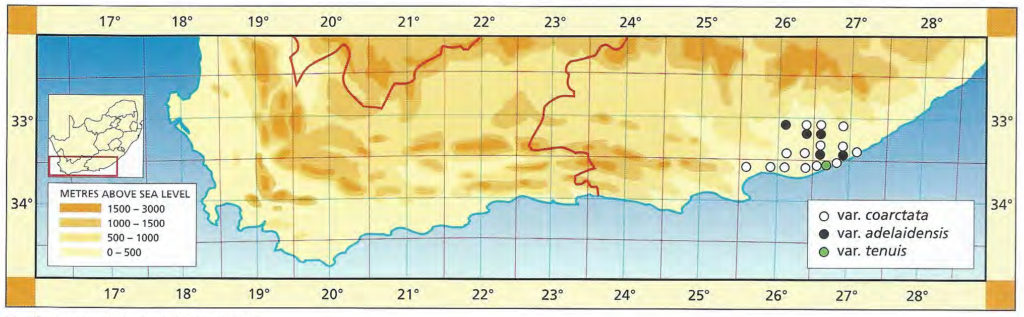
a. var. mutica. This is the element west of the Breede river. It was once known from northwest of Drew (north of the Breede River) from where it was collected and sold in pre-war times. That locality has since been taken in by wheat fields, and a single solitary clone survives in the Karoo Botanic garden collection. Leaves of this clone have been propagated. It is remarkable for the white opacity which has subsequently developed in the leaves, and which is also appearing in offsets of the original clone.
Distribution: 3320 (Montagu): N. Drew (-CC), Smith 5614 (NBG); Drew (-CC), Fouche in PRE 34916. 3419 (Caledon): E. Riviersonderend (-BB), Otzen 6 in Smith 5478 (NBG); 10km E. Riviersonderend (-BB), Smith 3270, 3460, 3970, 5478 (NBG), Bayer 4479 (NBG); Klipdale (-DB), (NBG). 3420 (Bredasdorp): E. Stormsvlei (-AA), Smith 3249 (NBG); Noukloof (-AA), Bruyns (NBG); Rietfontein (-AA), Bayer 2526 (NBG); Crodini (-AB), Bayer (NBG); Soesriver (-AC), Bayer 4473, in KG623/69 (NBG); Kykoedie (-AC), Bayer in KG83/77 (NBG); N. Kykoedie (-AC), Bayer in KG327/71 (NBG); Badjieskraal (-AC), Bayer in KG85/72 (NBG); Haarwegskloof (-AD) Venter 2 (NBG).
Inadequately located: ex hort, Malherbe, Smith 3430 (NBG); Caledon, Hurling & Neil (BOL); Bredasdorp, Theunissen (BOL).
Haworthia mutica var. mutica JDV92/64 Drew. The only clone known to have survived avaricious collecting. Collected in 1970, it has developed a curious and progressive white cloudiness in the leaves – also observed, albeit rarely in other collections.
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. This species flowers earlier than H. retusa.
Haworthia mutica var. mutica JDV85/17 northeast of Bredasdorp. In the field the plants have a purplish waxy bloom to the leaves.
b. var. nigra var. nov. Type: CAPE-3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG, Holo.).
nigra: black.
Differs from the type in its very dark coloration and its occurrence south and east of Heidelberg. (A var. mutica colore perfuscato differt).
This variety from Kransriviermond has been known for a long time and was distributed as H. retusa. It is very dark green, although this does depend on growing conditions as under softer light it may be quite green. There are also populations, probably of this variety, just east of Heidelberg.
Distribution: 3420 (Bredasdorp): Kransriviermond (-BB), Smith 5753 (NBG); Heidelberg (-BB), Smith 5509 (NBG); E. Heidelberg (-BB), Smith 5755 (NBG); 2km W. Heidelberg (-BB), Smith 6196, 6567 (NBG); N. Heidelberg (-BB), Smith 5509 (PRE).
Haworthia mutica var. nigra JDV86/16 south of Heidelberg. Less wax-like and dark green.
Haworthia mutica var. nigra JDV90/1 east of Heidelberg. There are three such populations known which seem to link H. mutica to H. turgida and to H. retusa.
25. Haworthia nortieri Smith, JS.Afr.Bot. 12:13(1946). Bayer :141(1976). Bayer :49(1982). Scott :88(1985). Type: Cape, Vanrhynsdorp, Smith 1676a (NBG): H. nortieri var. montana idem. 16:6(1950). Type: Cape, Clanwilliam, Smith 1678 (NBG): H. nortieri var. giftbergensis G.G.Sm. ibid. 16:7 (1950). Type: Cape, Vanrhynsdorp, Smith 7199 (NBG).
nortieri: for Dr. Nortier.
Rosette stemless, proliferous, 3-5cm φ. Leaves 25-45, soft sub-erect, ovate-lanceolate to obovate, pale to purplish green, with translucent spots on the leaves, small spines on margins and keel. Inflorescence slender, to 30cm. Flowers greyish-white, yellowish in tube.
1982 – H. nortieri occurs in the area between Clanwilliam and Vanrhynsdorp, extending inland to the foot of Vanrhyns Pass and on top of the plateau of the northern Cedarberg mountains. The var. globosiflora occurs in the dryer Botterkloof area and is distinguished by the flower having a globose tube while the leaves are shorter and broader. However, the flowers are not always so inflated and plants in the Vanrhyns Pass area have normal flowers while being vegetatively similar to the var. globosiflora. This is the basis for rejecting species status for the last-named. H. nortieri occupies the mid‑western geographic locale for the genus. The distribution southwards is not known and there is a big gap between this species at Pakhuis Pass (near Clanwilliam) and a form of H. archeri var. dimorpha in the Elandskloof area southeast of Citrusdal. H. arachnoidea occurs to the north of Vanrhynsdorp and also to the north and far west of Vanrhyns Pass. It does not, however, seem to appear in the Botterkloof area. H. nortieri is distinguished largely by the opaque leaf surfaces with abrupt, ovoid, pellucid spots. The flowers, and particularly the buds, are greyish in colour but the colour inside the tube is variable.
1999 – The range of H. nortieri has been extended considerably. The reference to H. archeri var. dimorpha at Elandskloof is quite erroneous and simply arises from the difficulty in relating that collection to the nearest known species. On the otherhand, the Elandskloof plants are only known to me from herbarium record and from two living plants collected by Drs Muller-Doblies. These did not seem to unequivocally be H. nortieri. P.V. Bruyns has collected H. nortieri from as far north as the Groenriver, to south as far as Kromriver in the Cedarberg. The late Harry Hall also collected it in the northeastern Knersvlakte. The most southerly collection is from near Opdieberg, north of Ceres. These plants also resemble the var. globosiflora but the flowers are not globose. At both extremes the plants tend to resemble H. globosiflora vegetatively . The plants at Komkans tend to have globose florets, and this is also true of plants at Groenriver where the florets are short and squat. The decision to include H. pehlemanniae within this species is also on account of the flower which is identical to that of var. globosiflora. This is not only in shape, but in colour too. Although the flowers may be the usual white with greenish veins, brownish-green flowers have been observed in both elements. The reference in the original description to quadrantly as opposed to spirally arranged flowers defies plant growth principles. The distribution of the two elements is complementary and gives the species as a whole an extraordinary cosmopolitan character. The colour in the flowers of the typical variety can be remarkable and as if yellow paint had been daubed at the throat of the florets. Habitat ranges from the moist south slopes of the Cedarberg to the dry wastes of Namaqualand and the so-called Moordenaarskaroo.
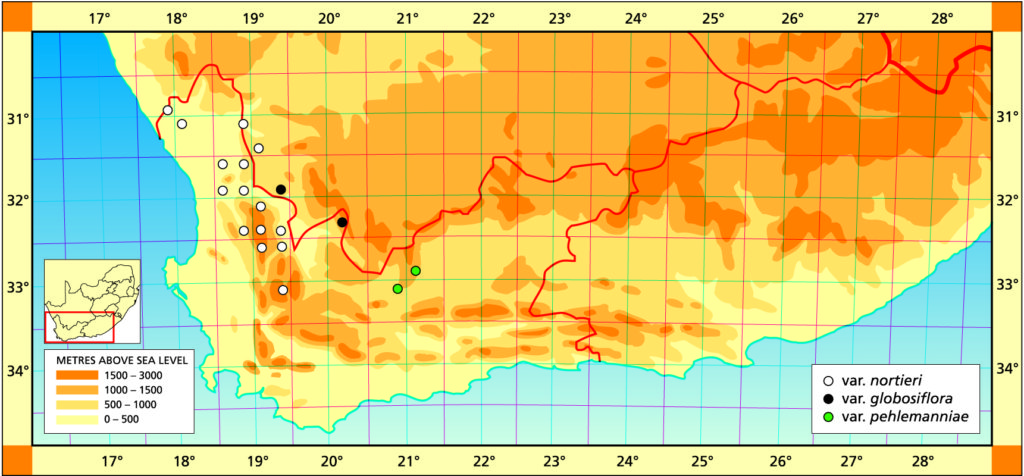
a. var. nortieri The typical variety occurs over a very wide area in the Table Mountain sandstones from south-east of Citrusdal to south-west of Nieuwoudtville. In addition it extends out into the Namaqualand lowlands, into very arid conditions. It is thus a very variable taxon in its own right.
Distribution: 3017 (Hondeklipbaai): Groenriver (-DD), Bruyns 6728 (NBG). 3118(Vanrhynsdorp): Komkans (-AA), Bruyns 6146 (NBG); Klipdrif (-BB), Hall 3390 (NBG); Gifberg (-DA), Smith 7199 (BOL,NBG), Thomas in NBG626/69; Steenkampskop (-DB), Bruyns 6167 (NBG); Kobe Pass (-DB), Bruyns 6170 (NBG); Die Kom (-DC), Bayer in KG329/72 (NBG); Doornriver (-DC), Smith 1676 (NBG), Leighton (BOL); W. Doornriver (-DC), Bayer 3637 (NBG); SE. Klawer (-DC), Leipoldt 4146 (BOL), Herre in STE6695 (BOL); E. Doornriver (-DD), Smith 1676a (BOL), Smith 6212 (BOL, NBG), Esterhuysen 6008 (BOL). 3119 (Calvinia): Uitkomst (-AC), Barker 10753 (NBG); Vanrhyns Pass (-AC), Smith 6211 (NBG), Hall in NBG656/60 (NBG), Ross-Frames in NBG1200/26 (BOL). 3218 (Clanwilliam): S. Clanwilliam (-BD), Van Jaarsveld 8153 (NBG). 3219 (Wuppertal): Pakhuis Pass (-AA), Smith 1678 (NBG), ex hort Whitehill NBG68397; Waboomsriver (-AC), Henderson 2212 (NBG); Diamond Drift (-AC), Leipoldt 3107 (BOL); N. Dwarsrivier (-AC), Bruyns in Bayer 6505 (NBG); E. Dwarsriver (-AD), Bruyns in Bayer 6506 (NBG); Heksberge (-CA), Smith 6116 (NBG), Muller-Doblies 79/015 (NBG), E. Elandskloof (-CA), Esterhuysen 3987 (BOL); Sandfontein (-CB), Esterhuysen 27199a (BOL); Cedarberg (-CB), Wagener in NBG11/43 (NBG); Nuwerus (-CB), Bruyns (NBG). 3319(Worcester): NW. Ceres (-AB), Aslander 645 (NBG).
Haworthia nortieri var. nortieri JDV93/61 Pakhuis Pass. Ocuring as a solitary specimen, as seen here, or in clusters.
Haworthia nortieri var. nortieri PVB6728 Groenriver. In this collection the flowers are very short but not globose.
Haworthia nortieri var. nortieri JDV88/34 Klawer. A variety of the Western Cape sandstones – not penetrated by H. Arachnoidea.
Haworthia nortieri var. nortieri JDV88/34 Klawer. The flowers are normal for the subgenus, but usually have a bright yellow throat.
b. var. globosiflora (Smith) Bayer :119(1976). Bayer 49(1982). H. globosiflora Smith, JS.Afr.Bot. 16:11(1950). Scott :87(1985). Type: CAPE‑3119 (Calvinia): Doornbosch, N. Doorn River Bridge (‑CD), Smith 7198 (NBG).
globosiflora: rounded flowers.
This variety is not known from only the Doornbosch area south of Botterkloof, which is also relatively unexplored. It has also been collected from as far east as the Ouberg Pass, southwest of Sutherland. The illustration in Scott (:88, 1985) is not of this variety at all and is probably of H. decipiens.
Distribution: 3119 (Calvinia): Doornbosch, (‑CD), Smith 7198 (NBG); 50km N. Clanwilliam (-CD), Dyer 3750 (PRE); Botterkloof (-CD), Hall in NBG68414, Villet (BOL); Boontjiesrivier, Kansekraal (-CD), Leipoldt 4119 (BOL). 3220 (Sutherland): Ouberg Pass (-AC), Venter (NBG).
Haworthia nortieri var. globosiflora JDV93/62 Botterkloof northwest of Clanwilliam. Typically with globose flowers, there are identical forms with ordinary flowers. In the winter they often “relax” so far that they lie flat on the soil.
Haworthia nortieri var. globosiflora JDV88/40 Ouberg Pass.This variety is in more karoid dry vegetation.
c. var. pehlemanniae (Scott) Bayer comb. nov. H. pehlemanniae Scott, Cact.Succ.J.(U.S.) 54:70(1982). Scott :79(1085). Type: CAPE-3320 (Laingsburg): 5km W. of Laingsburg (-BB), Scott 7450 (PRE).
pehlemanniae: for Inge Pehlemann.
Since first collected, this variety has been found at several other localities in the close vicinity of Laingsburg, but also further north in the Moordenaarskaroo and north of Matjesfontein. It differs from the species in the absence of the translucent spots on the leaf, and the vegetative similarity to H. arachnoidea in the same area is deceptive. It does appear to favour shales in relatively exposed situations as opposed to H. arachnoidea which generally prefers cooler south slopes. The two taxa grow in very close association.
Distribution: 3221 (Merweville): Klipfontein, N. Laingsburg (-CC), Aslander 801 (NBG). 3320(Montagu): 5km SW. Laingsburg (-BB), Scott 7450 (PRE), Bayer 3906 (NBG); N. Laingsburg (-BB), Venter (NBG).
Haworthia nortieri var. pehlemanniae JDV91/47 north of Laingsburg. Southwest of Laingsburg it occurs in among small sandstone boulders, whereas elsewhere it is in shales.
Haworthia nortieri var. pehlemanniae JDV87/204 southwest of Laingsburg. Although its leaves lack the transclucent spots of var. globosiflora, the flowers are identical.
Haworthia nortieri var. pehlemanniae JDV91/47 north of Laingsburg. Probably overlooked because of its similarity to H. arachnoidea.
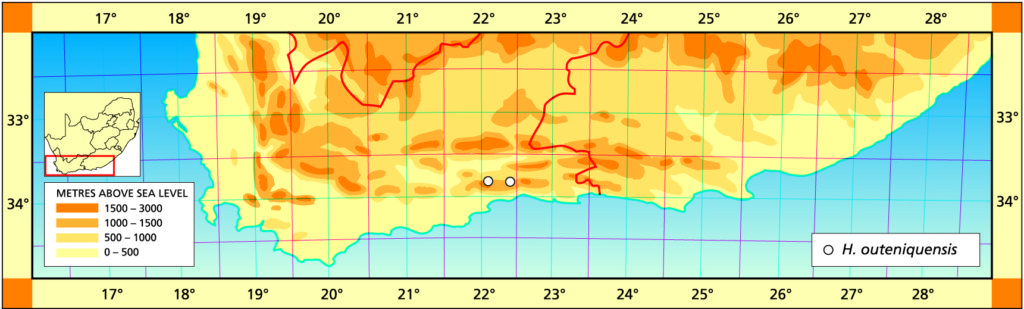
The form of this species is similar to that of H. monticola, but the difference is that the leaves are pellucid to translucent and firmer in texture. In brighter light the pellucid markings become more translucent and the opaque venation becomes reddish in colour. The margins of the leaves are quite cartilaginous. The distribution is complementary to that of H. monticola and H. chloracantha and one would have hoped that such populations strengthened the view of continuity between those two species. However, this does not seem to be the case. The two known populations are some considerable distance apart but in both cases are in exposed situations in sandstone in dry Fynbos. Venter suggests that the flowers relate to H. magnifica. (H. monticoleis affinis sed foliis solidioribus flavo-viridibus venatione rebella et marginibus carilagineis differt.)
Rosette acaulescent, 40-60mm diam., proliferous by off-set. Stem up to 15mm diam. Leaves erect to sub-erect, up to 60 X 6-10mm, 2-3mm thick, incurving tips with 2mm long end-awn, surfaces with pellucid anastomising dots, yellowish-green; face convex with 4-5 prominent rows of conspicuous pellucid dots; back convex with 3-6 rows of pellucid dots, with sharper keel bearing spines to 1mm long; margins similarly toothed, margins and keel pellucid. Peduncle simple, 1-1.5mm diam., 300-450mm long including raceme, brownish-green; base smooth, 2-angled; raceme with 7-15 flowers, 1-3 open; pedicels 4-6mm long, 1mm diam., sterile bracts 10-14, 8-10mm long. Perianth white and yellowish-green, 15mm long, perigon sub-triangular, tepals slightly bilabiate, upper outer tepals slightly replicate; tube yellowish-green within; buds slightly biarcuate, with slightly flattened pink tips. Flowering December.
27. Haworthia parksiana V.Poelln., Cactus J 5:34(1936). ibid., Feddes Repert.Spec.Nov. 41:205(1937). ibid. 43:104(1938). ibid., Desert.Pl.Life 10:48(1938). Bayer :143 (1976). Bayer :50 (1982). Scott :121 (1985). Type: Cape, Great Brak River, Mrs Helm in PE Parks 636/32. Not preserved. Lectotype (B&M). icon (B).
parksiana: in honour of ‘Mrs Parks’.
Rosette stemless, proliferous, 3-4cm φ. Leaves 25-35, 1,5-3cm long, blackish-green, sharply recurved, minutely tubercled, tip barely pointed. Inflorescence simple, slender, to 20cm. Flowers few, narrow, whitish with dull greenish venation.
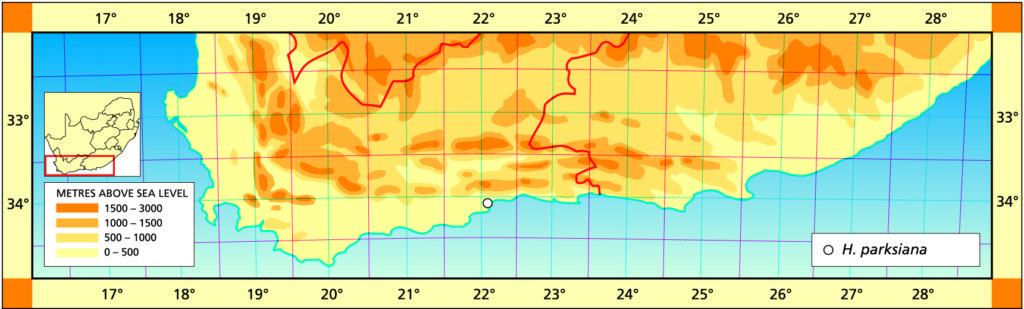
1982 – H. parksiana acquired its name in a very odd fashion. Von Poellnitz received the plants from F.R. Long under a Port Elizabeth Parks and Recreation Department collecting number ‘Parks 636/32’. Long had in turn received the plants from Mrs Helm of Great Brak. Von Poellnitz in error attributed the collection to a ‘Mrs Parks’ and hence the name. It is a very distinctive and very small dark species occurring very locally in Mossel Bay and the Great Brak area. Rather than having the flattened retused end area of the typical ‘retused’ species, the leaves are recurved at the ends. The flower buds are also round‑tipped at the end. The most obvious relationship that H. parksiana has with other species is with H. floribunda, although these two species are separated by the Gouritz River valley. H. parksiana, apart from being the smallest species, is also possibly the rarest and occurs only in very small numbers in the wild. It grows completely under the protection of small karroid shrubs, well hidden in the fallen leaf debris or among moss and lichen. The Mossel Bay area receives rain throughout the year but H. parksiana is particular about not having too much water. Offsetting does occur but growth is very slow.
1999 – The species is known to me from 3 localities. One of these is within the township of Great Brak, the other is on a rocky spur jutting out into a district road, and the third is on the very edge of a wheat field. Its continued existence thus seems to be fairly precarious. The relationship is with H. floribunda which is known (in terms of its probable or possible relation to H. chloracantha) at its most eastward, north of Albertinia. That population apparently includes forms which are comparable with H. parksiana. J.D. Venter cites the case where he has grown field collected seed of H. magnifica from south of Riversdale and observed parksiana-like plants among the offspring. This is what should be expected in terms of the reciprocity of forms, and also in terms of a ‘chaos’-driven speciation theory. Otherwise the species is readily recognisable and a relief from the contorted synonymies of other species.
Distribution: 3422 (Mossel Bay): Dumbie Dykes (-AA), Bayer 157 (NBG); Great Brak (-AA), James in NBG8106/45 (BOL), de Wildt (BOL), Smith 2936 (NBG); Botteliersberg (-AA), Bayer (NBG).
Haworthia parksiana JDV94/2 Great Brak. There is not much variation in this species but there is a reason to speculate continuity with H. floribunda.
Haworthia parksiana JDV96/87 Little Brak. This was a dense population among lichens on a small rock outcrop.
Haworthia parksiana JDV94/2 Great Brak. Can cluster profusely, yet still be very cryptic under dry conditions.
Rosette stemless, seldom proliferous, to 4cm φ. Leaves 20-35, short incurved, opaque grey-green, covered with minute spines. Inflorescence simple, to 20cm. Flowers 10-15, with upper lobes flared, white with pinkish venation.
1982 – H. pubescens has the same growth form as H. herbacea but it is smaller and seldom exceeds 30mm in diameter. The incurved leaves are dark grey‑green and finely pubescent. The flowers are quite unlike those of H. magnifica to which it may have been related, as the buds are long and slender. Also the upper perianth lobes are widespread as in H. herbacea. It grows in very close association with H. herbacea and flowers in November/December ‑ after H. herbacea and before H. magnifica. The distribution is extremely limited and only occurs on two low quartzitic hills east of the locality for H. maculata. It appears to be represented again about 15 km south by a form which is less pubescent and with more turgid leaves. This latter form at Lemoenpoort is in a quartzitic ridge again near H. herbacea, and it resembles H. maculata in the relative proportions of its leaves. H. maculata occurs in a more recognisable form about 5 km to the west at Moddergat, at a far southern locality for the species.
1999 – The geographic scale of species and their distribution ranges changes from the summer rainfall areas of the country to the southwestern winter rainfall area. H. pubescens probably has to be seen in that context. There seems to be little doubt that it is filling the space of H. maraisii, but that could also be mooted for H. maculata. The former species is the most probable relative as there are growth forms with the same leaf texture, and rosette shape near to Robertson. However, H. maraisii occurs in its typical form very close to Lemoenpoort at Trappieskraalkloof, just to the east. G.J. Payne did inform me that he had collected a small dark species just above the Brandvlei Prison and this would probably fall into this context if it is re-collected.
Haworthia pubescens var. pubescens JDV84/7 south of Worcester. Very seldom growing in exposed situations.
Haworthia pubescens var. pubescens JDV84/7 south of Worcester. An elegant localized species with affinities with H. maraisii. Apart from the growth form it also flowers in late spring and has a flower more similar to H. herbacea.
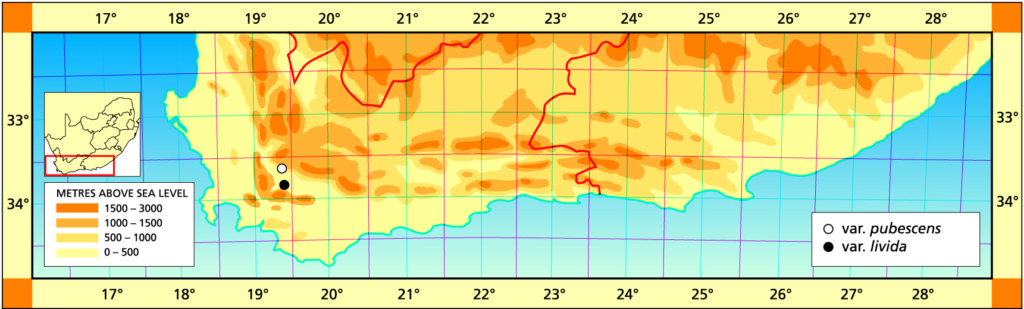
b. var. livida var.nov. Type: CAPE-3319(Worcester): S. Lemoenpoort (-CD), Bayer 1128 (NBG, Holo.).
livida: bluish-grey.
Differs from the typical variety in being less pubescent, with slightly broader and fewer leaves, and partly with pellucid spots on the leaves. (A var. pubescens foliis latioribus minus pubescentibus cum maculis pellucidis differt).
Distribution: 3319 (Worcester): S. Lemoenpoort (-CD), Bayer 1128 (NBG).
Haworthia pubescens var. livida JDV88/31 Lemoenpoort. Close examination suggests similarities with species as unlikely as H. marumiana.
Haworthia pubescens var. livida JDV88/31 Lemoenpoort, south of Worcester. Broader leaves and sparsely pubescent.
[ed.] Bayer now considers var. livida as H. maculata var. livida
H. maculata var. livida (M. B. Bayer) M. B. Bayer (Haworthia Nomenclator, 10, 2012). Type: RSA, Western Cape (Bayer 1128 [NBG]). — Distr: RSA (Western Cape: Robertson Karoo: S of Worcester). ≡ Haworthia pubescens var. livida M. B. Bayer (1999) ≡ Haworthia maraisii var. livida (M. B. Bayer) M. Hayashi (2000) ≡ Haworthia intermedia var. livida (M. B. Bayer) J. Esterhuizen (2003); incl.Haworthia livida Breuer (2011) (nom. inval., ICN Art. 38.1a, 41.5). Differs from var. maculata: Ros smaller; L less spotted.
Rosette stemless, occasionally proliferous, to 5cm φ. Leaves 30-45, incurved, coriaceous, block-patterned reticulation, dark- to emerald-green, with pronounced whitish spines on margins and keel. Inflorescence simple, slender, to 30cm. Flowers 15-20, white.
1982 – This is a small compact, dark‑green species. The leaves are armed with pronounced white spines. The distribution is from near Constable Station in the east, to west and south of Touws River. Von Poellnitz recorded H. angustifolia from Touws River but it is difficult to believe that he was confusing this species with H. pulchella. The leaves in the latter species are firmer, shorter and incurved. H. pulchella is always in the shade and is also a winter‑growing species. It is very seldom proliferous and is very slow growing. The most likely relative is probably H. wittebergensis primarily on the basis of distribution and the moderately coriaceous texture of the leaves.
1999 – The range of this species has also been extended with new records from further to the east, at least as far as the Anysberg. There is a will to associate this species with H. wittebergensis which I think overlooks the more probable affinity of that species to H. blackburniae on the basis of stem structure (granular and woody) and on the same amplexicaul leaf bases. H. pulchella has a soft textured stem and the leaves are normally inserted on the stem as for, say, H. marumiana. This comparison is made because Col Scott suggests the extension of H. pulchella up to the Nuweveld Mountains at Beaufort West where he suggests that it also occurs, although his distribution map does not show this. That is incorrect as H. pulchella is with fair certainty restricted to the Touws River-Laingsburg-Hoekvandieberg triangle. It is curious to see the resemblance which H. maraisii has to this species growing on identical Witteberg quartzites, together even with Protea sulphurea and other Dry Mountain Fynbos species, near Bonnievale. The distribution of H. margaritifera supports this distant connection and it may be more reasonable to seek some affinity of H. pulchella in the south-west, rather than eastward. The species is not restricted to shade and specimens have been seen growing in very exposed sites, although obviously not preferred.
Haworthia pulchella var. pulchella JDV87/187 southeast of Touws River. Usually solitary, there is at least one population in which the plants form large clusters.
Haworthia pulchella var. pulchella JDV87/187 southeast of Touws River. It is not easy to grow, possibly because of its winter rainfall distribution.
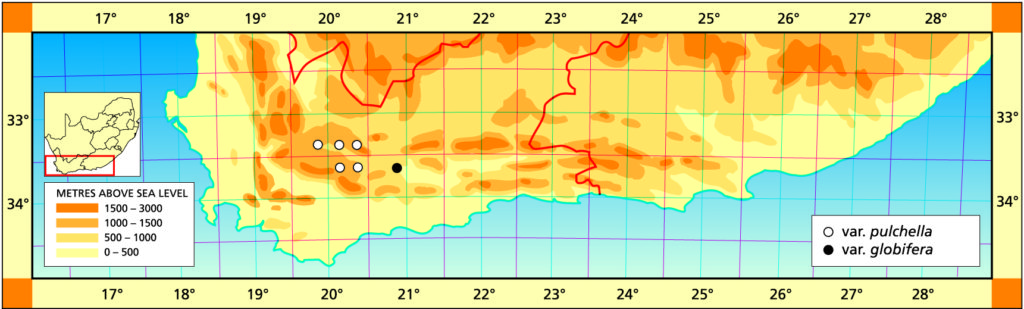
b. var. globifera var. nov. Type: CAPE-3320 (Montagu): SE Anysberg Bruyns 7338 (BOL, Holo.).
globifera: forming globose clusters and rosettes.
Differs from the species in being glabrous, slightly stem forming and also forming clusters. (A var. pulchella foliis glabris differt.)
The typical variety is seldom proliferous and usually slowly so. The population southwest of the Anysberg is unusual in that the plants form compact clusters well protected on south facing slopes. The new variety requires less protection, forms larger, more raised clusters and there is some degree of stem formation with offsets above the base. The leaves are incurved to form very neat, tight clusters which suggested the name.
Distribution: 3319 (Montagu): SE Anysberg (-DB), Bruyns 7338 (BOL).
Haworthia pulchella var. globifera JDV97/147 Touwsberg. A glabrous, clustering and slightly stem forming variety. Unlike the typical variety, it occurs on very hot north slopes. The leaves are pale yellowish-green in winter, rich red in summer.
30. Haworthia pygmaea V.Poelln., Feddes Repert.Spec.Nov. 27:132 (1930). ibid. 41:208 (1937). ibid. 43:104 (1938). ibid., Kakteenkunde 9:104 (1937). Bayer :148:(1976). Bayer :51 (1982). Type: Cape, Great Brak, Mrs van der Bijl. Not preserved. Neotype (B&M)): E. Great Brak, Fourcade 4759 (BOL). H. asperula Haw. p.p. sensu C.L.Scott :119 (1985).
pygmaea: dwarf.
Rosette stemless, slowly proliferous, 6-10cm φ. Leaves 12-15, retused, round-tipped, surface pellucid with obscure raised tubercles, sometimes intensely papillose. Inflorescence simple, robust, to 30cm. Flowers white with greenish veins.
1982 – The name is not very apt as H. pygmaea is not much smaller than any of its near relatives. In fact it is very much bigger than H. parksiana, with which it grows. However, in the field some plants only develop 2‑3 leaves and perhaps von Poellnitz received some of these. The species is characterised by scabrid or obscurely papillate to papillate leaf surfaces, the leaf end-areas are flattish and the leaf tips rounded. Von Poellnitz ascribed plants from Great Brak to several other species too, and there is a close similarity with some forms of H. emelyae, H. retusa and H. mutica. Geographical considerations weigh heavily in retaining H. pygmaea as a species separate from H. retusa, as it is apparently confined to the area between Mossel Bay and Great Brak. H. retusa var. dekenahii occurs east of Albertinia, approaching Mossel Bay, and perhaps could be regarded as intermediate. H. turgida changes fairly dramatically as it moves eastward and it occurs very close to H. pygmaea. Thus these two species may be related in the same way that H. turgida is related to H. retusa.
1999 – The previous discussion is inaccurate in several respects. The forms of H. emelyae referred to were probably of H. bayeri. The forms of H. retusa were those tending to H. magnifica. H. retusa var. dekenahii east of Albertinia is of course actually the var. argenteo-maculosa of H. dekenahii Smith, which in this work is regarded as a variety of H. pygmaea. The recognition of the relationship between H. retusa and H. turgida negates the possibility of a similar relationship between H. pygmaea and H. turgida. The form that has received most of the attention is the very shiny papillate one. It does not occur in any specified area or as a distinctive population and varietal rank is not warranted. There is a difficulty in distinguishing H. pygmaea from H. mutica in cultivation but the former should be recognisable by the presence of the surface tubercles. These are however, very much less conspicuous than in H. magnifica var. splendens.
a. var. pygmaea. Confined to the Great Brak and Mossel Bay area.
Distribution: 3422 (Mossel Bay): Great Brak (‑AA), Bayer 2241 (NBG), Morris in PRE 34890, Luckhoff in NBG1871/24 (BOL); Near Great Brak (-AA), Luckhoff 6269 (PRE); E. Great Brak (-AA), Fourcade 4759 (BOL); W. Great Brak (-AA), Smith 2919 (NBG), Bayer 2287 (NBG); Dumbie Dykes (-AA), Bayer 2289 (NBG).
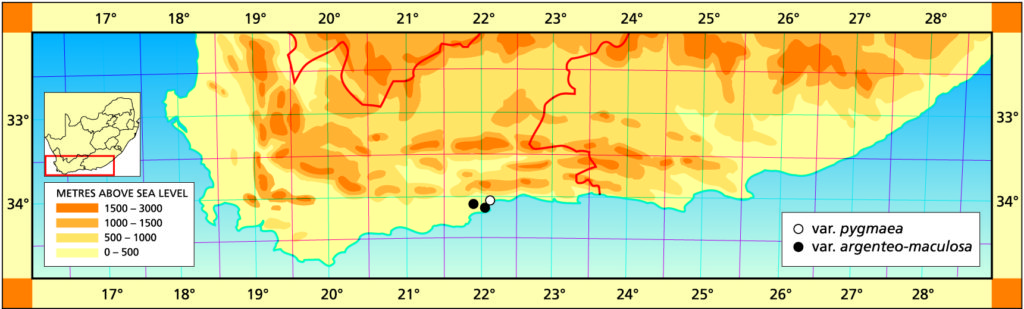
b. var. argenteo-maculosa (Smith) Bayer comb. nov. Bayer, Aloe 34: 6(1997), H. dekenahii var. argenteo-maculosa Smith, J.S.Afr.Bot. 11:74 (1945). H. retusa forma argenteo-maculosa (Smith) Bayer :98(1976). Type: CAPE‑3421 (Riversdale): between Gouritz Bridge and Mossel Bay (‑BB), S. Emett in NBG 68037.
argenteo-maculosa: silver spotted.
The locality for this variety could be at one of at least two localities known east of the Gouritz Bridge and the name is also applied to a population just west of Mossel Bay itself. There is undoubtedly a very strong link with H. magnifica var. splendens which has more conspicuous surface tubercles. The variety is separated from the typical species by the more conspicuous white flecking in the leaves and also by its relative smoothness.
Distribution: 3421 (Riversdale): between Gouritz Bridge and Mossel Bay (‑BB), S. Emett in NBG 68037; A few km E. Gouritz River Bridge (-BB), Smith 3959 (NBG,PRE); Cooper Siding (-BB), Bayer (NBG); Humor (-BB), Bayer (NBG). 3422 (Mossel Bay): W. Mossel Bay (-AA), Schoemann (NBG).
Haworthia pygmaea var. argenteo-maculosa JDV93/65 west of Mossel Bay. The population reveals the connection to the H. magnifica complex.
Haworthia pygmaea var. argenteo-maculosa JDV91/38 Humor. The white flecks can be very pronounced.
Haworthia pygmaea var. argenteo-maculosa JDV91/38 Humor. The separation from H. magnifica may be dubious as it is only a question of the less conspicuous surface tubercles.
Haworthia pygmaea var. argenteo-maculosa JDV86/75 Cooper Siding. The similarity of the populations shown on this page makes it very difficult to classify them rationally.
31. Haworthia reticulata Haw., Syn.Pl.Succ. :94(1812). Haw., Rev.Pl.Succ. :57(1921). V.Poelln., Feddes Repert.Spec.Nov. 44:233(1938). Bayer, Natn.Cact.Succ.J 27:10(1972). Bayer :150(1976). Bayer :52(1982). Aloe reticulata Haw., Trans.Linn.Soc. 7:9(1804). A. arachnoidea var. reticulata (Haw.) Ker-Gawl., Curtis’ Bot.Mag. :t1314. Type: Cape. Not preserved. Neotype: Icon. Curtis’ Bot.Mag. :t1314(1811): Aloe pumilio Jacq., Hort. Schoenbr. 4:11(1804). Type: icon, Hort. Schoenbr.: H. reticulata var. acuminata idem. 43:93(1938). Type: Cape, Robertson, G.J. Payne. Not preserved. Neotype (designated here): CAPE-3319(Worcester): Kliphoogte, SW. Robertson (-DD), Smith 3986 (NBG): H. hurlingii var. ambigua Triebn. et V.Poelln. idem. 43:93(1938). Type: Cape, Montagu, G.J. Payne in Triebn. 906. Not preserved. Neotype (designated here): CAPE-3319(Worcester): Wolfkloof, Robertson (-DD), Bayer 1543 (NBG): H. guttata Uitew. in Desert Pl.Life 19:136(1947). Type: Cape, Robertson to Bonnievale. Not preserved.
reticulata: like a network.
Rosette proliferous, partially stemmed, to 8cm φ. Leaves 25-40, firm sub-erect, incurved, lanceolate-acuminate, opaque with reticulate to mottled patterning, margins and keel frequently with short spines, reddening in sun. Inflorescence simple, to 25cm. Flowers large, white to pinkish, with arcuate buds with flattened tips.
1982 – H. reticulata is closely allied with H. herbacea. It occurs between Worcester and Robertson generally north of the Breede River. In the area southwest of Robertson it occurs also south of the river and it is sometimes difficult to determine if the species is H. reticulata or H. herbacea. H. reticulata extends along the Breede River to about Drew and is not known further east. At Bonnievale the plants have narrower more denticulate leaves. The distinction between this species and H. herbacea is that it is generally more glabrous, more proliferous and grows higher above the ground. The flowers are usually pinkish as opposed to beige in H. herbacea. The var. hurlingii occurs between Robertson and Bonnievale and has short obtuse leaves. It is small and seldom exceeds 20‑30mm in diameter. H. reticulata is very much bigger in the lower Hex River Valley where it hybridises with H. herbacea. Nearby, the two species are again adjacent but do not hybridise. H. reticulata is very much more variable than H. herbacea and it is possible to see an affinity with H. turgida ‑ especially with the Swellendam and Bredasdorp populations of that species. Deeply spotted forms of H. reticulata occur in the Eilandia area southwest of Robertson, and it is probable that Uitewaal’s H. guttata was a form of H. reticulata after all.
1999 – This species is now well-known and is very common in the Worcester-Robertson area. Von Poellnitz three times seemed to confuse specimens from the Eastern Cape with this species. He described H. haageana in 1930 and in 1937 the var. subreticulata. In the first case with H. haageana, it is certain that the origin was just incorrectly cited as Grahamstown. Regarding H. haageana var. subreticulata he may in fact have had a plant which did not belong with H. reticulata at all, although he also later conceded that perhaps the cited origin at Grahamstown was perhaps incorrect. In the case of H. reticulata the flower should always be diagnostic. The species does vary considerably over its range and there have been several varieties described to cover this. The varieties upheld here should be seen as a communication about variation as a process of revision, rather than expressing an absolute division of the species. Ultimately communication may only be effective at the level of geographic locality and origin.
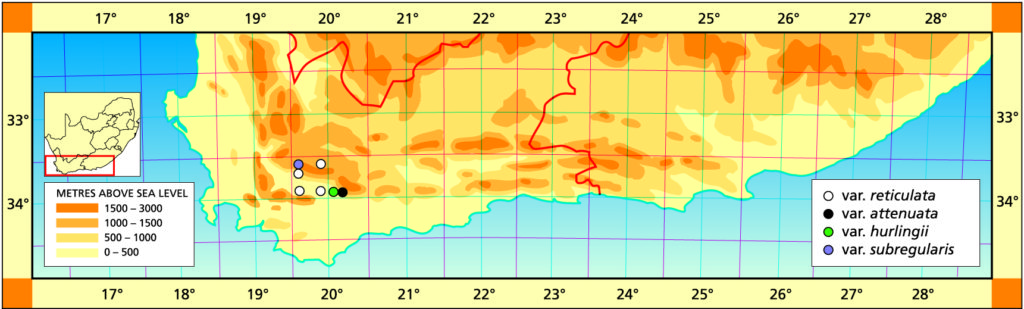
a. var. reticulata. There are several populations which are very close in appearance to H. herbacea and the difference is usually that the latter species does not readily form dense clumps and the flower is more buff-coloured. The close relationship of the two species is a very good indication of how the species in the genus generally can relate to one another. Each population is fairly distinctive and the varieties formalised here are only a sample of this quite extensive variation, which is often associated with the intergradation with H. herbacea.
Distribution: 3319 (Worcester): 24km NE. Worcester (-DA), Bayliss 2198 (PRE); Keeromskloof (-DA), Bayer in KG662/69 (NBG); Buitenstekloof (-DB), Bayer (NBG); Rooikleigat (-DC), Bayer 1438 (NBG); Ribbokkop (-DC), Bayer 160 (NBG); S. Gemsbokkop (-DC), Bayer 1539 (NBG); Dublin (-DC), Bayer (NBG); Rooiberg (-DC), Bayer in KG175/70, in KG90/76 (NBG); Kliphoogte, SW. Robertson (-DD), Smith 3986 (NBG); Wolfkloof, Robertson (-DD), Smith 3983 (NBG), Bayer 1543 (NBG, PRE), Bayer 2187, 4665 (NBG); Olifantshoogte (-DD), Hurling & Neil (BOL).
Inadequately located: ex hort, Rosch & LeRoux 596 (PRE); Hurling & Neil in BOL24592, van der Merwe 172 (BOL)
Haworthia reticulata var. reticulata JDV86/107 Buitenstekloof. Occurs in very large clusters on a dolomitic outcrop. H. intermedia V. Poelln. was in the earlier Handbook mistakenly believed to have referred to this population.
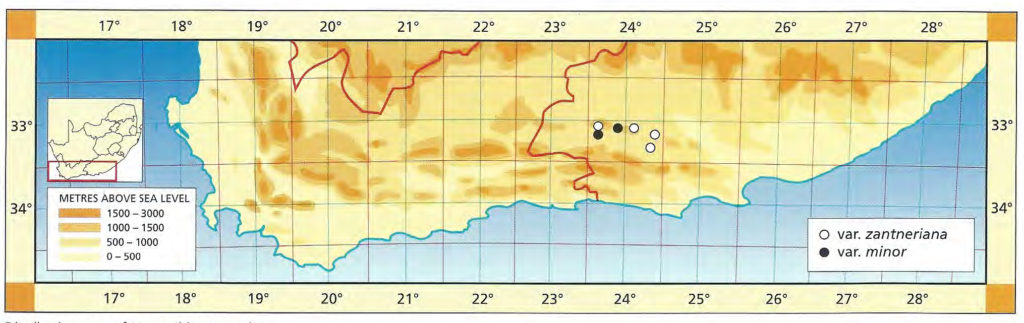
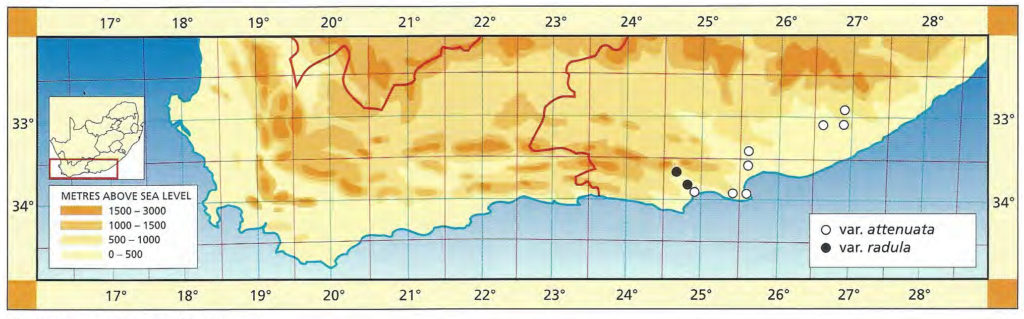
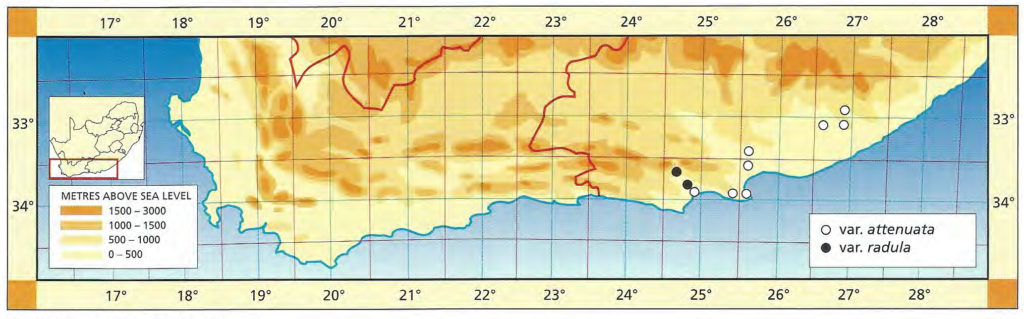
b. var. attenuata var. nov. Type: CAPE-3320 (Montagu): 2km S. Bonnievale, Smith 3979 (NBG, Holo.).
attenuata: with leaves narrowing to a point.
Differs from the species in having longer, more slender leaves. (A var. reticulata foliis longioribus gracilioribusque differt).
Although there is a plethora of names already applied to variants of the species, this variety is described to emphasise the southeastern range of the species in the Bonnievale/Drew area. The plants have more attenuate leaves and they are also more robustly spined. Curiously there is a resemblance to H. variegata var. modesta, particularly in those clones with relatively flat unmarked upper leaf surfaces and weak spination.
Distribution: 3320 (Montagu): 2km S. Bonnievale (-CC), Smith 3979 (NBG); Bonnievale (-CC), J. Smith 6283 (PRE), van der Merwe (BOL); Angora (-CC), Hurling & Neil (BOL); 5km SE. Bonnievale (-CC), Bayer 4665 (NBG).
Haworthia reticulata var. attenuata JDV90/28 south of Bonnieville. The marginal spines are quite pronounced in this variety.
Haworthia reticulata var. attenuata JDV91/36 southwest of Bonnieville. Here the plants have less attenuate leaves than further eastward.
c. var. hurlingii (V.Poelln.) Bayer :52(1982). H. hurlingii V.Poelln., Cactus J 5:34(1936). V.Poelln., Feddes Repert.Spec.Nov. 41:202(1937). V.Poelln. Cactus J 6:19(1937). V.Poelln., Desert Pl.Life 10:125(1938). Type: Cape, from Bonnievale, Mr Hurling, Stellenbosch. Not preserved. Neotype (B&M): icon (B).
hurlingii: for Hurling.
Von Poellnitz suggests that this variety grows deeply buried in the ground. This is not true for the species nor for this variety, which forms small clumps. It is characterised by its small obtuse leaves and compact shape. There are other populations in which the individual rosettes are even smaller and fairly densely spined e.g. Bosluiskloof, southwest of Robertson.
Distribution: 3320(Montagu): Near Bonnievale (‑CC), Smith 3445 (NBG); 18km E. Bonnievale (-CC), Fouche 64 (PRE); Bonnievale (-CC), Hurling & Neil 6283b (BOL,PRE), Hurling & Neil in NBG2341/35, Smith 3442 (NBG); Goudmyn (-CC), Bayer 4659, in KG89/76 (NBG), (NBG); 7km W. Bonnievale (-CC), Smith 3257, 3981 (NBG).
Inadequately located: ex hort Malherbe in NBG482/42, Marais in NBG1241/36, Smith 3274, 3893 (NBG).
Haworthia reticulata var. hurlingii JDV87/207 southwest of Robertson. This is the type locality for H. hurlingii var. ambigua which is regarded here as a variant of the typical variety.
Haworthia reticulata var. hurlingii JDV87/191 west of Bonnievale. There is a second population further to the west.
Haworthia reticulata var. hurlingii JDV87/191 west of Bonnievale. Much smaller rosettes.
d. var. subregularis (Bak.) Bayer comb. nov. H. subregularis Baker, Saund.Ref.Bot. 4:t232(1870). Baker, J Linn.Soc.Bot. 18:391(1880). Baker, Fl.Cap. 6:212(1896). V.Poelln. Cactus J 5:236(1936). Type: Cape without locality, Copper. Not preserved. Lectotype (designated here): Icon. Saund.Ref.Bot. :t232: H. haageana V.Poelln., Feddes Repert.Spec.Nov. 28:104(1930). Type: Cape, imported from Grahamstown. Not preserved: H. haageana var. subreticulata V.Poelln., Cact. J 5:37(1937). idem. Cact.J 6:18(1938). idem. Feddes Rep.Spec.Nov. 44:232(1938). Type: Cape, Grahamstown, Mrs Ferguson. Not preserved.
subregularis: almost regular.
In the 1982 edition, mention was made of the larger size of plants from the ‘lower’ Hex River Valley. The reference is specifically to the DeWet area northeast of Worcester where the plants are indeed large, with often spreading leaves as in the Ref.Bot. illustration. Baker names the plant for the regularity of the perianth lobes but he is undoubtedly using the same free licence that other botanists have used in comparing flowers of Haworthia with Astroloba, or indeed the flowers of the Haworthia subgenera. In my opinion he noticed the distinctiveness of the flower respective to others of the subgenus without true cognition of what the differences really were.
Distribution: 3319 (Worcester): De Wet (-DA), Van Breda 223 (PRE), Malherbe in NBG478/42, Barker 516 (NBG), Hurling & Neil in NBG542/35, Stayner (NBG), Smith 3232, 3443 (NBG), Peers (BOL); Hex River Pass (-DA), Barker 7455 (NBG); Boskloof (-DA), Bayer in KG324/71 (NBG); Tweefontein (-DA), Smith 7387 (NBG).
Inadequately located : ex Whitehill, NBG68287, Venter in NBG142/39, Venter 2 (BOL), Meiring in NBG 4538/14 (BOL), Smith 3242, 3353, 5091, 5157, 7269 (NBG).
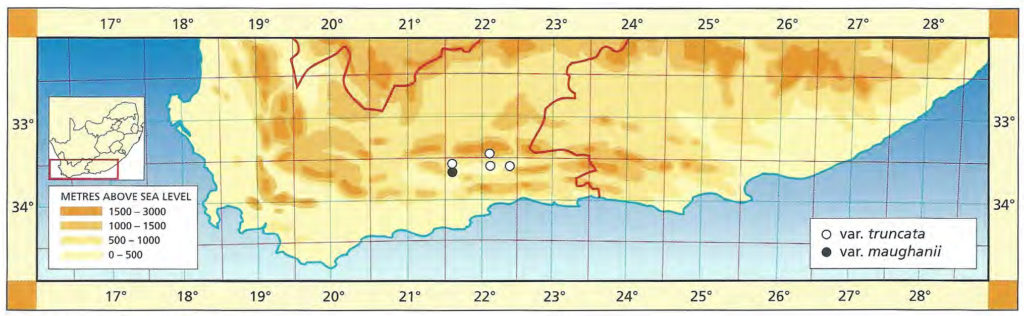
32. Haworthia retusa (L.) Duval, Pl.Succ.Hort.Alenc. :7(1809). Haw., Syn.Pl.Succ. :95(1812). Bayer :150(1976). Bayer in Excelsa 8:46(1979). Bayer :53(1982). pp Scott :112(1986). Aloe retusa L. Sp.Pl. :322(1753). Haw., Trans.Linn.Soc. 7:9(1804). Type: icon, 2:t6 Commelin, Hort.Amstel.(1701): H. foucheii V.Poelln., Succulenta 22:28(1940). Type: Cape, Riversdale district, Grootvlei, Fouche. Not preserved. Neotype (designated here): CAPE-3421(Riversdale): Near Riversdale, Grootvlei (-AB), Dekenah 212 (NBG): H. retusa var. multilineata Smith, JS.Afr.Bot. 12:3(1946). H. multilineata (Smith) Scott :135(1985). Type: CAPE-3421(Riversdale): 3km N. Riversdale, J. Dekenah 83 in Smith 5383 (NBG): H. retusa var. solitaria Smith, JS.Afr.Bot. 12:5(1946). H. solitaria (Smith) Scott, Aloe 11:37(1973). Type (designated here): CAPE-3421(Riversdale): along Corrente River, 10km N Riversdale, Dekenah 5 in Smith 5373 (NBG): H. retusa var. densiflora Smith, JS.Afr.Bot. 12:7(1946). Type: CAPE-3421(Riversdale): Riversdale district, Venter 106 in Smith 5056 (NBG): H. geraldii Scott, JS.Afr.Bot. 31:123(1965). Scott, Aloe 11:22(1973). Scott :132(1985). Type: CAPE-3421(Riversdale): 5km east of Riversdale, Scott 72 (PRE).3
retusa: with leaf-tips bent back thumb-like.
Rosette stemless, slowly or seldom proliferous, to 12cm φ. Leaves 10-15, turgid, rigid, with pronounced retused end-area, pointed tips, variously lined and windowed. Without surface spination and usually without spination on margins and keel. Colours brownish or green and seldom purpling. Inflorescence simple, robust, to 30cm. Flowers compacted on inflorescence, white with greenish-brown veins.
1982 – H. retusa is very closely associated with H. turgida, and an eventual re-evaluation of this association could lead to a total upheaval of species concepts in this group of haworthias. H. retusa is taken to be an assemblage of forms in the Heidelberg and Riversdale areas, with clearly defined end areas, and points, to the leaves. The various forms may or may not be proliferous, for example the var. solitaria was largely solitary, whereas the form described as H. geraldii is very proliferous. The forms vary in cultivation and both dark and light green forms may occur. The darker forms may have some relationship to H. magnifica, and the light forms to H. turgida. The species is not well known in the area between Heidelberg and the Breede River and hence it is not known whether or not H. mutica and H. retusa intergrade. H. mutica may simply be a blunt‑leaved form of H. retusa. The var. dekenahii from Albertinia is now thought to be represented by several populations in which plants have leaves very silver‑spotted ‑ a phenomenon which occurs in H. turgida too. This variety may eventually be shown to link H. retusa and H. pygmaea. There is still an apparent break in the distribution of these two species between Mossel Bay and Albertinia. The var. acuminata tends to be darker green and the leaves are very acuminate. It occurs in the south‑east of the distribution range of the species and may also be expected in the area southwest of Albertinia ‑ still unexplored.
1999 – This revised treatment fulfils the predictions of 1982 amid the realisation that the views expressed there are correct, but stopping short of uniting H. retusa with H. turgida. H. longebracteata Smith is now here regarded as a variant of that ubiquitous species and strictly H. geraldii Scott should be treated similarly. However, the key issue is that H. retusa, as perceived here, is the robust, generally solitary forms which occur only in the Riversdale area. Scott’s treatment is rather fortuitous in that he typifies the name in the same way, but applies it to the smaller clump-forming elements which comprise H turgida. This is evident from his synonymy but totally compromised in his discussion where:- 1. he says it is restricted to one locality at Riversdale and two to three in the Little Karoo, and 2. the distribution map where localities at Little Brak and at Tradouw pass seem to be indicated. Breuer and Metzing’s argument for the creation of an epitype, and also their choice, is also unfortunate. The Commelin illustration could hardly be more distinctive and apart from Col Scott who muddled H. retusa and H. turgida, there has never ever been any sign of doubt about its application. Furthermore, they select a specimen from the source of H. fouche. The Commelin, illustration depicts the acute leaf-tips of the species very well and there is little chance of confusion with H. mutica
In the species concept for the genus, co-occurrence and consequent interaction, or lack thereof, are criteria for recognising species. In the field it has become obvious that the interaction between species revolves around H. turgida as a main role-player and not H. retusa. There is no interaction between the latter two named elements as they do not co-occur. The three species recognised by Scott viz. geraldii, fouchei and multilineata give a very good impression of the variation of this one species in the Riversdale area. The var. solitaria may represent interaction with H. magnifica, and the vars. acuminata and dekenahii are both transferred to that species. H. dekenahii var. argenteo-maculosa is treated as a variety of H. pygmaea. The problem plants around Heidelberg are associated with H. mutica.
Distribution: 3421 (Riversdale): 8km W. Riversdale (-AA), Smith 5443 (NBG); Along Corrente River, 10km NW. Riversdale (-AA), Dekenah 5 in Smith 5373 (NBG), Dekenah in KG305/71 (NBG); 4km N. Riversadel (-AA), Smith 5493 (NBG); SW. Riversdale (-AA), Smith 5387 (NBG); Near Riversdale, Grootvlei (-AB), Dekenah 212 (NBG), Smith 4955 (NBG), Fourcade 263 (NBG), Bayer in KG627/69 (NBG), Bohnen 9058 (NBG); Blinkbonnie (-AB), Smith 6086 (NBG), Venter in KG 156/71; Zeekoegat (-AB), Smith 6089 (NBG); Ferguson Drive (-AB), Smith 5380 (NBG, PRE); 3km N. Riversdale (-AB), J. Dekenah 83 in Smith 5383 (NBG), Smith 5374, 5488 (NBG, PRE), Smith 6796 (NBG); 5km E. Riversdale (-AB), Scott 72 (PRE), Smith 5377 (NBG); Vet River Road (-AB), Smith 5383, 5387 (PRE); 14km E. Riversdale (-AB), Smith 5749, 7320 (NBG); Bolus 2868 (PRE); Commonage (-AB), Smith 5377 (PRE); Dekenah 11 (PRE); Riversdale (-AB), Fouche in PRE 34869; 3km E., Grootvlei (-AB), J. Scott 1778 (PRE); 3km E. (-AB), J. Scott 93 (PRE);
Inadequately located: Riversdale district, Venter 106 in Smith 5056, Smith 3432, 3850, 3919, 3962, 5043, 5056, 5599 (NBG), Bolus 11390 (BOL), Helm in NBG1746/32, Malherbe in NBG399/40, ex hort, Wilman in PRE 34904.
Haworthia retusa JDV93/57 east of Riversdale. Some forms are reminiscent of H. magnifica.
Haworthia retusa JDV89/26 south of Riversdale. Others are enormous and range from brown to yellow.
Haworthia retusa JDV92/122 southeast of Riversdale. A motly assemblage where some are conspicuously veined.
Haworthia retusa JDV92/122 southeast of Riversdale. Some clones remain very compact.
Haworthia retusa JDV92/122 southeast of Riversdale. Despite the size there is an attempt at concealment.
Haworthia retusa JDV92/69 southeast of Riversdale. A cultivar referred to as ‘Jolly Green Giant’ probably originated in this area.
Haworthia retusa JDV87/5 east of Riversdale. The well known fa. geraldii, which is not accepted in this treatement although the name could be retained for practical and historical reasons.
Haworthia retusa JDV87/5 east of Riversdale. This form is only known from one small site where it forms massive clusters.
Haworthia retusa JDV92/125 east of Riversdale. This is the fa. fouchei, occuring a stone’s throw from geraldii but also not upheld for the same reasons.
Haworthia retusa JDV92/125 east of Riversdale. As with H. magnifica var. magnifica, there are two populations of this H. retusa variety on the same ridge. This is also the fa. fouchei, recognized by its erect leabes.
Haworthia retusa JDV92/125 east of Riversdale. The leaves are more erect and there is continuity down the Kafferkuilsrivier with H. turgida.
[ed. Bayer subsequently resurrected some of the varieties.]
H. retusa (Linné) Duval (Pl. Succ. Horto Alencon., 7, 1809). Type: [lecto — icono]: Commelin, Horti Med. Amstelod. 2: 11, t. 6, 1701. — Distr: RSA (Western Cape); Fynbos vegetation.
H. retusa var. longibracteata (G. G. Smith) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type: RSA, Western Cape ( Dekenah 18 in Smith 5378 [NBG, PRE]). — Distr: RSA (Western Cape). I: Scott ( 1985: 127, as H. longibracteata).
≡ Haworthia longibracteata G. G. Smith (1945) ≡ Haworthia retusa fa. longibracteata (G. G. Smith) Pilbeam (1983) ≡ Haworthia turgida var. longibracteata (G. G. Smith) M. B. Bayer ( 1999).
Differs from var. retusa: L erect to suberect, ovate-lanceolate.
H. retusa var. nigra (M. B. Bayer) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type: RSA, Western Cape ( Smith 5753 [NBG]). — Lit: Bayer ( 2004a); Bayer ( 2004c); Bayer ( 2005); all as H. mutica var. Distr: RSA (Western Cape: Heidelberg area).
≡ Haworthia mutica var. nigra M. B. Bayer ( 1999) ≡ Haworthia silviae var. nigra (M. B. Bayer) M. Hayashi (2000); incl.Haworthia chromutica M. Hayashi (2000) ( nom. inval., ICN Art. 39.1, 40.1); incl.Haworthia quimutica Breuer ( 2011) ( nom. inval., ICN Art. 38.1a).
Differs from var. retusa: Ros more proliferous; L greener and more translucent.
H. retusa var. retusa — Distr: RSA (Western Cape: Riversdale). I: Bayer ( 1982: Fig. 33a).
Incl.Haworthia fouchei Von Poellnitz (1940) ≡ Haworthia retusa fa. fouchei (Von Poellnitz) Pilbeam (1983) ≡ Haworthia retusa var. fouchei (Von Poellnitz) Breuer (2016); incl.Haworthia retusa var. densiflora G. G. Smith (1946); incl.Haworthia retusa var. multilineata G. G. Smith (1946) ≡ Haworthia retusa fa. multilineata (G. G. Smith) Pilbeam (1983) ≡ Haworthia multilineata (G. G. Smith) C. L. Scott (1985); incl.Haworthia retusa var. solitaria G. G. Smith (1946) ≡ Haworthia solitaria (G. G. Smith) C. L. Scott (1973); incl.Haworthia geraldii C. L. Scott (1965) ≡ Haworthia retusa fa. geraldii (C. L. Scott) Pilbeam (1983) ( nom. inval., ICN Art. 41.5); incl.Haworthia retusa var. quimutica Hayashi (2001); incl.Haworthia subretusa Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a).
Ros stemless, rarely slowly proliferating, to 12 cm ∅; L 10–15, turgid, rigid, with pronouncedly retuse end-areas, 8 × 2 cm, brownish or green and rarely with purplish hue, variously lined and windowed, surface and usually also margins and keel without spines or tubercles, tips pointed; Inf robust, to 30 cm; Fl 20–30, closely spaced, white with greenish-brown veins.
Possible hybridization with H. mirabilis is mentioned by Bayer ( 2012d).
H. retusa var. suberecta (Von Poellnitz) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type [neo]: RSA, Western Cape ( Bayer s.n. in Karoo Garden 631/69 [NBG]). — Distr: RSA (Western Cape). I: Scott ( 1985: 126, as H. dekenahii).
≡Haworthia turgida var. suberecta Von Poellnitz (1938) ≡ Haworthia turgida fa. suberecta (Von Poellnitz) Pilbeam (1983) ≡ Haworthia suberecta (Poellnitz) Breuer ( 2010); incl.Haworthia turgida var. subtuberculata Von Poellnitz (1938); incl.Haworthia turgida var. pallidifolia G. G. Smith (1946) ≡ Haworthia turgida fa. pallidifolia (G. G. Smith) Pilbeam (1983) ≡ Haworthia pallidifolia (G. G. Smith) M. Hayashi (2010) ≡ Haworthia suberecta var. pallidifolia (G. G. Smith) Breuer (2016); incl.Haworthia pseuda Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a); incl.Haworthia reflexa Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a); incl.Haworthia rodinii Breuer ( 2010) ( nom. inval., ICN Art. 36.1b, 38.1a).
Differs from var. retusa: L strongly mottled, tips slightly truncate and rounded.
H. retusa var. turgida (Haworth) M. B. Bayer (Haworthia Update Vol. 7, Part 4, 36, 2012). Type [neo]: RSA, Western Cape ( Bayer 2420 [NBG 132378]). — Distr: RSA (Western Cape). I: Bayer ( 1982: Fig. 40, as H. turgida).
Differs from var. turgida: Ros partially stemless, proliferous, 5–10 cm ∅; L 20–40, ovate-lanceolate, 4 × 1.2 cm, turgid, often as thick as broad, recurved or slightly retuse, margins and keel lightly spined.
33. Haworthia semiviva (V.Poelln.) Bayer :153(1976). Bayer :54(1982). Scott :89(1985). H. bolusii var. semiviva V.Poelln., Feddes Repert.Spec.Nov. 44:135(1938). Type: Cape, Beaufort West, Marais in Triebn. 1080. Not preserved. Lectotype (B&M)): icon (B).
semiviva: half alive.
Rosette stemless, seldom proliferous, 5-6cm φ. Leaves 30-40, broad ovate, thin, incurved, translucent and usually necrotic at tips. Inflorescence simple, 20-30cm. Flowers white with green venation, broad across tube.
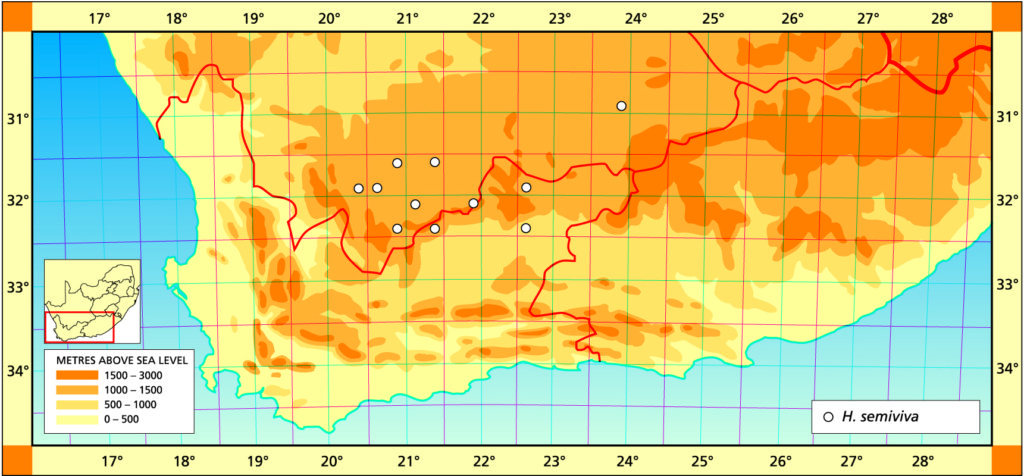
1982 – H. semiviva is not well‑known, but it occurs as far west as Middelpos and may skirt the northern margins of the Nuweveld mountains to appear again at and just south of Beaufort West. Originally this species was described as a variety of H. bolusii but this is only a partly true reflection of the relationship as is borne out by the distribution. H. semiviva is not known east of Beaufort West and there is thus a break in distribution between the two species. Unlike H. bolusii, the tips of the leaves die well back so that the plants are protected by the whitened leaf ends as in H. lockwoodii. In cultivation the leaf-tips are flattened and highly translucent; which is a tendency, as in H. lockwoodii, to the absence of any pigmentation at the ends of the leaves. H. semiviva does not proliferate and is only occasionally found as a 2‑ or 3‑headed plant. It is difficult and slow in cultivation. Although also technically in the summer rainfall region, it is a winter‑growing species. Like other species it also grows in the protection of small karoo shrubs in accumulated windblown sand and organic debris.
1999 – It appears that this species was first collected at the southeastern most part of its range as it is now well known from the Frazerburg, Sutherland, Victoria East area. It may transpose to H. bolusii var. blackbeardiana in the far northwest which would become evident in the retention of live end-area to the leaves. Although it is tempting to suggest an affinity with H. lockwoodii, there has been no evidence of populations which link them geographically, and neither do they share the bluish-green colour of the cooperi/bolusii/decipiens var. cyanea alliance. Where the two species might be expected to meet in the central lower Karoo, one finds H. decipiens var. cyanea.
Haworthia semiviva JDV93/47 north of Three Sisters. More like the typical form but the leaf tips are barely dry and white.
Haworthia semiviva JDV97/60 Beaufort West. The typical form has no keel to the leaves and the tips are dried white.
Haworthia semiviva JDV97/18 south of Victoria West. This is possibly a transition to H. bolusii var. blackbeardiana. It is a very white frosted looking specimen but the leaves are keeled, uncharacteristic for H. semivia.
34. Haworthia serrata Bayer, JS.Afr.Bot. 39:249(1973). Bayer :55(1976). non Scott :62(1985). Type: CAPE‑3420 (Bredasdorp): Oudekraalkop, Heidelberg (‑BA), Bayer 166 (NBG).
serrata: toothed.
Rosette stemless, seldom proliferous, to 7cm φ. Leaves 20-30, narrow, acuminate, spined along margins and keel, bright yellowish-green with translucent lines above. Inflorescence simple, robust, to 40cm. Flowers white with green venation.
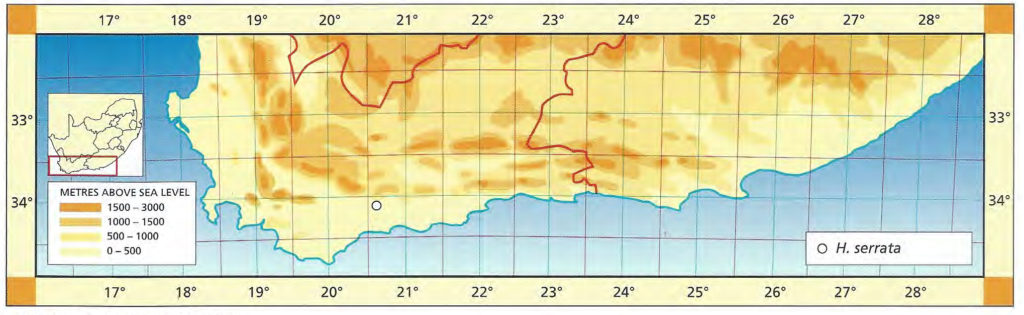
1982 – H. serrata occurs in an area which appears to be a kind of watershed for the genus. This is the area between the Duiwenhoks river draining from Heidelberg to the sea, and the Breede River. Neither H. retusa, H. turgida, nor H. magnifica are known in the area, and only the one variant regarded as a variant of H. heidelbergensis is known 20 km southwest of Heidelberg at Matjestoon. H. serrata has the same general form as H. herbacea but is a more uniformly green plant with larger leaves which have a tendency to bend outward. The flowers are large and white, unlike H. retusa and H. turgida, and more closely resemble those of H. emelyae from north of the Langeberg mountains. Recent collections by C. Burgers from the coastal limestones may throw more light on the relationship of H. serrata with a population at Bredasdorp and H. heidelbergensis, and more collecting needs to be done immediately east of the Breede River too.
1999 – The 1982 discussion is vague regarding the Bredasdorp connection. However, this is because the Bredasdorp area, and particularly the coastal limestones, seem to harbour some unusual elements (see H. mirabilis var. calcarea, H. variegata var. petrophila and H. heidelbergensis var. minor). The reference was specifically to the last named taxon, which is in any case now recognised in this work. H. serrata can curiously best be understood by recognition of non-co-occurrences. There are no other species in the immediate localities known for this species and hence continuance is the only guide. In this perspective it can be seen to relate to both H. heidelbergensis and to H. mirabilis. Seedlings often include weaker forms which resemble either the Bredasdorp or Matjestoon varieties of H. heidelbergensis. Some plants of the Riversdale to Heidelberg populations of that species in turn bear resemblance to H. serrata. There is also a very close resemblance to forms of H. mirabilis var. sublineata. The flower is rather robust for the southern Cape species and thus mention was made of the similarity to H. emelyae var. multifolia. That element also has a connection to H. mirabilis probably through H. heidelbergensis. H. serrata occurs at three discrete localities and is virtually the only Haworthia species in that area between the Breede River and Heidelberg. The general shape of the plant tends to deviate from the retuse-like forms of its close allies and veers towards H. herbacea in that respect. Indeed this is the kind of variability one should expect in terms of Vavilov’s homology or Chaos theory. Col. Scott mistakenly took a form of H. mucronata for this species.
Haworthia serraata JDV87/148 southwest of Heidelberg. The flowers are large and very white in comparison with its neighbours.
Haworthia serraata JDV88/53 southwest of Heidelberg. Many fairly slender spreading leaves which maintain a bright green colour.
[ed.] Bayer now calls this species H. rossouwii V.Poelln. in Kakteenk. 7: 75 (1938).
H. rossouwii var. rossouwii — Distr: RSA (Western Cape: SW of Heidelberg to Bredasdorp); mainly in Fynbos vegetation. I: Bayer (1982: Fig. 36, as H. serrata). Incl.Haworthia serrata M. B. Bayer (1973) ≡ Haworthia chloracantha var. serrata (M. B. Bayer) Halda (1997) ≡ Haworthia rossouwii var. serrata (M. B. Bayer) Breuer (2016). Ros stemless, rarely proliferating, to 7 cm ∅; L 20–30, narrow, 6 × 1 cm, acuminate, bright yellowish-green with translucent lines above, margins and keel spiny; Inf robust, to 40 cm; Fl 20–30, white with green venation.
Rosette stemless, non-proliferous, to 10cm φ. Leaves 8-12, turgid, very rounded and retused, with translucent end-area and several longitudinal short lines. Inflorescence simple. Flowers white with brownish venation.
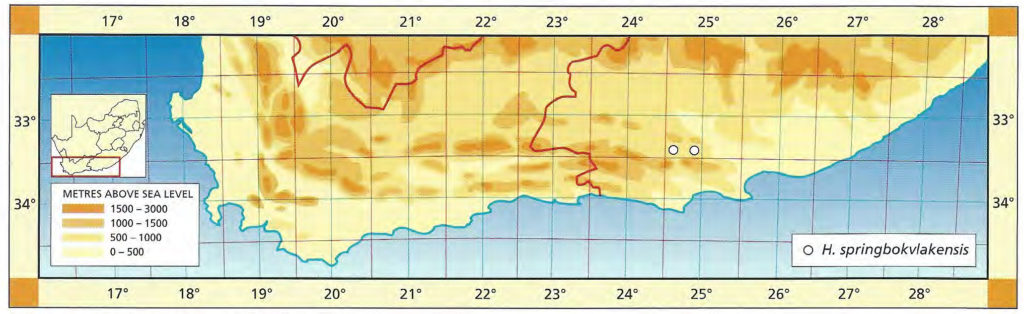
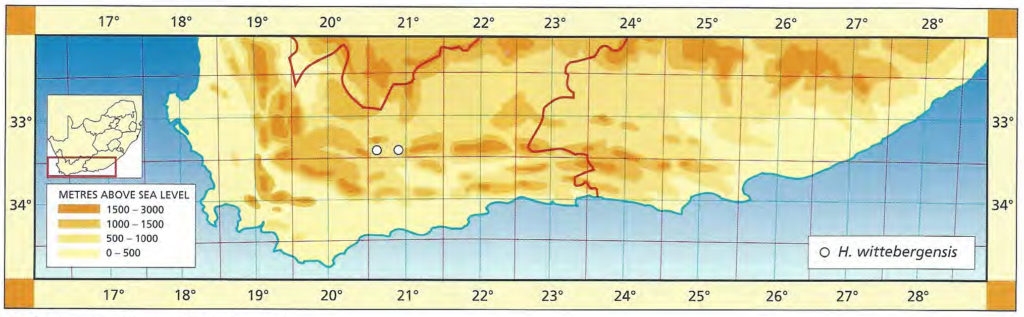
1982 – This very distinct species with very turgid leaf faces and rounded leaf tips is at the extreme east of the distribution range for the ‘retuse’ haworthias. H. emelyae would be supposed to be its nearest relative but this species is known only as far east as Uniondale. It seems as though the gap can only be explained by poor collecting. H. springbokvlakensis is not well‑known although there are reports of other localities in the area of the type. The plants are quite variable and the leaves tend to be slightly viscid so that clay and dust particles adhere tightly to leaf surfaces. Field collected plants are thus often unsightly and dirty compared to the well‑grown cultivated specimens. The relationship of H. springbokvlakensis to the newly discovered H. bruynsii in the subgenus Hexangulares is the most intriguing mystery in Haworthia.
1999 – The distribution of this species has not really been extended by any new collections apart from a collection reported west of Kirkwood by P. Schoemann and another in the Klipplaat area by J. Bouwer. The latter collection was of a single robust plant and there is no further record from this area. Mr A. Joubert reported the presence of the species on three different farms about the known type locality. There cannot be a relation with H. bruynsii as suggested in 1982 and perhaps the clarification of H. emelyae and H. bayeri will lend more credence to the probability that it is related to the latter. The species includes forms with very attractive reticulate venation in the upper exposed leaf surfaces which normally only have longitudinal venation.
Distribution: 3324 (Steytlerville): Springbokvlakte (-BD), Scott 245 (PRE), Smith 3585 (NBG), Bayer in KG402/70 (NBG); W. Kleinpoort (-BD), Smith 3590 (NBG); Kleinpoort (-BD), Smith 3585, 3238 (NBG); Two Waters (-BD), Smith 2916 (NBG).
Inadequately located: Malherbe in NBG1003/39, Barker 5068 (NBG), Smith 3530 (NBG).
Haworthia sprinbokvlakensis JDV96/98 east of Springbokvlakte. Some forms are almost without surface markings.
Haworthia sprinbokvlakensis JDV93/21 south of Kleinpoort. Others may be strongly reticulated.
Haworthia sprinbokvlakensis JDV91/11 west of Springbokvlakte. It is a wonderful plant to grow with its extraordinary rounded, turgid leaves.
36. Haworthia truncata Schonland, Trans.R.Soc.S.Afr. 1:391(1910). V.Poelln., Feddes Repert.Spec.Nov. 27:136(1930). ibid. 41:214(1937). Hutchinson, Cact.Succ.J(U.S.) 23:99(1951). Scott, Natn.Cact.Succ.J 29:36(1967). Bayer :134(1976). Bayer :57(1982). Scott :138(1985). Marx, Aloe 33:18(1996). Type: Cape Colony, near Oudtshoorn, Miss G. Britten (K): H. truncata fa tenuis V.Poelln., Feddes Repert.Spec.Nov. 44:239(1938). H. truncata var. tenuis (V.Poelln.) Bayer, Haworthia Handbook: 161(1976). Scott :138(1985). Type: Cape, Oudtshoorn area. Not preserved. H. truncata fa crassa V.Poelln. loc. cit. Type: Cape, Oudtshoorn area. Not preserved. Lectotype (B&M) icon, Des. Pl. Life 19: 79(1947): H. truncata fa normalis V. Poelln. ibid. Type: Cape, Oudtshoorn area. Not preserved.
truncata: ending abruptly and square.
Rosette stemless, slowly proliferous, distichous. Leaves 10-12, from 12mm to 40mm wide, 3mm to 10mm thick, scabrid with minute tubercles, end-area abruptly truncate, sub-pellucid. Inflorescence simple, to 200mm. Flowers white with brownish veins.
1982 – Like H. maughanii, the leaves are very abruptly truncated, but they are arranged distichously. This unusual leaf form and arrangement has earned the species the colloquial name ‘perdetande’, meaning ‘horses teeth’. H. truncata is quite widespread although there are many indications of gross over collecting. It occurs from west of Oudtshoorn near De Rust, to just southwest of Calitzdorp. Although reasonably variable it has been observed that the fa tenuis does not necessarily retain its small size when grown in cultivation and recognition of such variety no longer seems necessary. H. truncata is easy to grow and propagates from leaf and from root ‑ in both cases it appears necessary for there to be some stem tissue as a source of leaf primordia.
1999 – The Japanese have exploited the range of variation in this species and have some extraordinary cultivars which seem to make the recognition of varieties really superfluous. The fact that crossing H. truncata and H. maughanii seems to produce intermediates and plants equivalent to both parents, suggests that the difference of distichous to multifarious, is a very simple genetic one. Marx (1996) has reported on the two elements growing together in a polyglot of intermediary forms, thereby confirming a view that they are really conspecific. This requires repetition of a comment on hybridisation. It is really irrational to speak of hybrids in the way that suggests the parents have come from different origins and are now in cross-pollination contact. Here we have two elements which must have sprung from a common source and never been isolated. What that source is may be a mystery, and the most probable in terms of geographic location and morphology is perhaps H. bayeri. A form has also been collected in the same general area that the var. tenuis occurs, which has surface hairs much like H. cooperi var. venusta from the Eastern Cape.
a.var. truncata Distributed from southwest of Calitzdorp to the De Rust area east of Oudtshoorn. This variety has the capacity to re-grow from roots although it is not certain if stem tissue is required to achieve this.
Distribution: 3321 (Ladismith): Blackburn Valley, Calitzdorp(‑DA), W.F.Barker 5340 (NBG); SW. Calitzdorp (-DA), Smith 2069 (NBG); W. Calitzdorp (-DA), Venter 13 (NBG). 3322 (Oudtshoorn): Schoemanshoek (-AC), Oddie in NBG82/20 (BOL); Volmoed (-CA), Venter 7 (NBG), Heunis 9 (NBG); Oudtshoorn graveyard (-CA), Heunis 7 (NBG); DeRust to Kamanassie (-CB), Smith 6915 (NBG); Vanwykskraal (-CB), Fourcade 213 (NBG), Smith 2068, 4007, 5621 (NBG), Venter 20 (NBG);
Inadequately located: Cape, Marloth 12732 (PRE); Calitzdorp, Blackburn (BOL); Oudtshoorn, Taylor (BOL); ex hort Whitehill, NBG68235, Malherbe in NBG422/40.
Haworthia truncata var. truncata JDV85/82 Vanwykskraal. A famous locality and the source of most cultivated specimens.
Haworthia truncata var. truncata JDV85/82 Vanwykskraal. The capacity to regenerate from roots may account for the resilience of the population.
Haworthia truncata var. truncata JDV92/98 east of Oudtshoorn. A remarkable torm with spinescent leaf surfaces.
Haworthia truncata var. truncata JDV85/84 west of Calitzdorp. In this area there are plants with very thick leaves.
b.var. maughanii (V.Poelln.) Fearn, Natn.Cact.Succ.J 21:28(1966). H. maughanii V.Poelln., Feddes Repert.Spec.Nov. 31:85(1932). ibid. 41:205(1937). ibid. 44:239(1938). Bayer :134(1976). Bayer :47(1982). Scott :139(1985). Type: Cape, near Calitzdorp, H. Maughan-Brown. Not preserved. Neotype (B&M): CAPE‑3321 (Ladismith): Calitzdorp, Malherbe in NBG307/40 (BOL).
maughanii: for H. Maughan‑Brown.
Rosette multifarious. Leaves round in cross-section.
1982 – There are two species with abruptly truncated leaves, H. maughanii and H. truncata. In the former species the leaves are multifarious whereas in H. truncata they remain distichous. Artificial cross‑pollination has yielded hybrids with shared parental characters. Geographically H. maughanii occupies a very distinctive small habitat within the western limits of the distribution of H. truncata. The relationship of the two species is thus a little unclear and there may be little reason for maintaining two species. Contrary to a suggestion by Hutchinson (1951), the relationship with H. retusa must be distant. The truncation of the leaves is apparent in the earliest leaves and is not brought about by outward flexure of the leaf tips as occurs in H. retusa.
1999 – This element is also remarkably variable and the truncated end-area may be translucent, opaque and variously veined as in the typical variety. The end-area margins may be scabrous and variously crispate or undulate. Also the number, shape, and size of the leaves varies enormously and parallels easily the same range that motivated von Poellnitz to recognize three major forms in H. truncata.
Distribution: 3321 (Ladismith): S. Calitzdorp (-DA), Fourcade 169 (NBG), Smith 2070, 6094, 6094a (NBG); Calitzdorp (‑DA), Mrs Taute in NBG 68778.
Inadequately located: Calitzdorp, Blackburn (BOL), Ross-Frames (BOL), Taylor (BOL), Oddie in NBG1559/32 (BOL); ex hort, Whitehill (NBG),Malherbe in NBG307/40; Cape, Marloth 12732 (PRE).
Haworthia truncata var. maughanii JDV84/36 south of Calitzdorp. The exciting variants bred in captivity make it surprising that ordinary specimens from the field are still traded unscrupulously.
Haworthia truncata var. maughanii JDV84/36 south of Calitzdorp. Differs from the var. truncata only in that the leaves are rotate and not distichous. The leaf shapes and markings are also highly variable, providing wonderful material for selection and breeding.
Haworthia truncata var. maughanii JDV94/10 south of Calitzdorp. Fortunately ostriches discourage access to the limited locality. This is teh population cited by Marx, 1996.
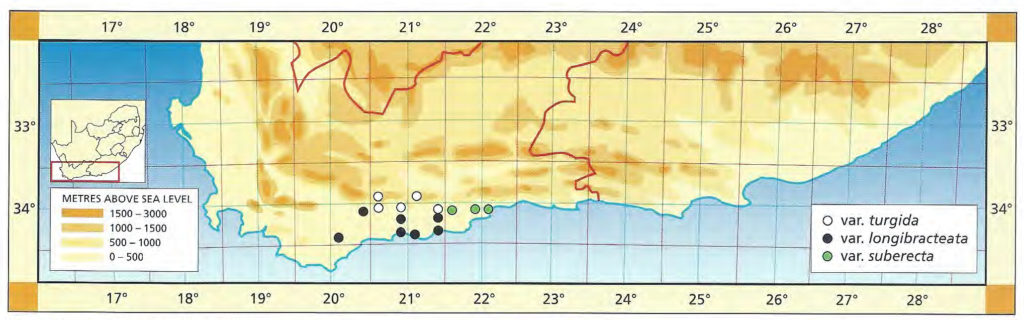
37. Haworthia turgida Haw., Suppl.Pl.Succ. :52(1819). Salm-Dyck, Monogr. 9:t5(1837). Bayer :163(1976). Bayer :57(1982). H. retusa sensu Scott :113(1985). Type: Cape, ex hort Kew. Not preserved. Neotype (B&M): icon, Bowie (K): H. laetivirens Haw., Suppl.Pl.Succ. :53(1819). Salm-Dyck, Monogr. 10:t3(1837). Type: Cape. Not preserved. Neotype (designated here): icon t.3, Salm-Dyck: H. caespitosa V.Poelln., Cactus J 5:33(1936). V.Poelln., Feddes Repert.Spec.Nov. 43:103(1938). Type: Cape, near McGregor, G.J. Payne in Triebn. 586. Not preserved. Lectotype (B&M): icon (B): H. caespitosa fa subplana V.Poelln. ibid. 44:232(1938). Type: Cape, Uniondale, G. Helm in Triebn. 874. Not preserved: H. caespitosa fa subproliferans V.Poelln. ibid. Type: Cape, Calvinia. Not preserved.
turgida: swollen.
Rosette partially stemless, proliferous, 5-10cm φ. Leaves 20-40, turgid often as thick as broad, recurved or slightly retused, generally mottled, yellow-green to pink in sun, margins and keel lightly spined. Inflorescence simple, 15-20cm. Flowers slender, brownish-white with darker venation.
1982 – H. turgida is the most widely distributed of the Southwestern Cape species and also the most variable. It occurs in both the higher mountains in sandstone, and also on the lower lying shales. It is very subject to ecotypic variation and the best evidence of this is north of Bredasdorp eastward apparently through the coastal limestones, northward to Swellendam where there is possibly intergradation with H. reticulata, then eastwards again in the Langeberg Mountains and down all the main river valleys including the lower Breede River valley. The eastern limit is near Little Brak. The northern limit is of course the Langeberg mountain range and it is unlikely that H. turgida occurs inland along the Gouritz river ‑ despite an unconfirmed report of a collection northwest of Calitzdorp. Some of Von Poellnitz’s forms and varieties were recorded from as far afield as Calvinia and Steytlerville. Although the possibility of such collections cannot be completely ruled out, it is unlikely that these could have been relatives of H. turgida. There are forms in the Potberg mountains as well as in the Riviersonderend mountains which may intergrade with other local species (e.g. at Greyton with H. mirabilis) in the same way that H. turgida is related to H. retusa in the Heidelberg and Riversdale areas, or at Albertinia. It is a fascinating problem as it is obvious that affinities at one locality may not be the same at another. Thus it is quite conceivable that H. turgida may have an affinity with H. magnifica var. notabilis at Robertson, with H. maculata in the Hex River valley and with H. archeri through the inland mountains. H. turgida is generally a small species up to about 30mm diameter in the sandstones, however, in shales it may grow up to 80mm diameter. It is always very proliferous and grows on steep rocky slopes as opposed to H. retusa which is less proliferous and on level areas. H. turgida is also not withdrawn into the ground. This contrast of proliferation and withdrawal into the soil as opposed to clump formation also occurs in the related species H. reticulata and H. herbacea, and is evident in less strongly related species such as H. cooperi and H. cymbiformis.
1999 – The early illustrations by Salm-Dyck hardly allow room for doubt about this species, and yet the localities that von Poellnitz cites are grossly off the mark for the species and varieties which he described and cited. The problem that Col Scott had with this species and with H. retusa should also be explained here. It arises out of a misconception about leaf tiers and the statement ‘quinquefarious’ in the early literature. This term refers to vertical leaf tiers and it is apparent from a manuscript in the Grahamstown (Albany) Museum (and also from the revision in Aloe 11, 1973) that Scott concluded this to mean horizontal leaf layers. Also Scott’s treatment, where he applies the name H. retusa to this element, as well as to a number of von Poellnitz’ species and varieties H. laetivirens, is baffling. Breuer and Metzing have nominated a neotype which is not a good representation of the typical variety from the sandstones of the Langeberg mountains.
Of the southern Cape species, H. turgida is unquestionably the main role player. It occurs in recognisable form from Bredasdorp to east of Mossel Bay. It forms a continuum with H. reticulata and H. herbacea which are in the Worcester/Robertson area, and ranges from the Langeberg high peaks to the valleys running seaward and southward. It is unfortunate that apart from Smith’s names, few others have any geographical credibility. The affinities suggested in the 1982 discussion are in retrospect improbable. If there is a further association which needs exploration, it may be the link with the inland species through the high mountain forms. It does not seem sound to speculate on the chronological origins of the elements at altitude as opposed to those at lower levels.
a.var. turgida The typical variety based on Salm-Dyck’s renditions, rather than on the Kew illustration cited by Scott, which I have not seen, is considered to be the one in the Tradouw Pass east of Swellendam. The leaves are almost recurved, highly mottled and moderately spined. The higher mountain forms in the sandstones are generally smaller and note has to be taken of similarities between these smaller softer elements of apparently disparate species e.g. H. mirabilis var. consanguinea, H. maculata, and H. vlokii, all at high altitude in sandstones. At Heidelberg H. turgida expresses its full potential in terms of ecotypic variation where it passes from sandstones, to shales and on to the clays of the Witteberg series. It also appears to hybridise with both H. heidelbergensis and with H. floribunda in that area.
Distribution: 3320 (Montagu): Tradouw Pass (-BC), Read (BOL), Smith 3247, 5160, 6784, 7517 (NBG); S. Barrydale (-DC), Smith 3902 (NBG). 3321(Ladismtih): 16km N. Riversdale (-CC), Smith 7197 (NBG); NE. Riversdale (-CC), Smith 5385 (NBG). 3420(Bredasdorp): Buffeljachts (-BA), Smith 4941 (NBG); Heidelberg (-BB), Smith 5044 (NBG), Kramer in NBG841/60; N. Heidelberg (-BB), Smith 6203 (NBG); Bayer in KG240/72, in KG 241/72 (NBG); Blackdown (-BB), Smith 5546 (NBG). 3421(Riversdale): Glen (-AB), Muir 3006 (BOL); Klein Kruisriver (-AB), Smith 6075, 6080 (NBG).
Inadequately located: ex hort. Whitehill, NBG68371, NBG68056, ex hort. Ross-Frames, NBG81/44, NBG2937/32, NBG68057, NBG1827/28, Smith 3240, 3429), 3498, 3904, 3950 (NBG); Swellendam, Ross-Frames (BOL); Riversdale, Muir in NBG1827/28 (BOL); Albertinia, Muir 1300 (BOL).
Haworthia turgida var. turgida JDV89/20 north of Heidelberg. This population seems to have been influenced by H. heidelbergensis.
Haworthia turgida var. turgida JDV89/23 north of Heidelberg. A little further north the plants are transforming to the smaller typical appearance.
Haworthia turgida var. turgida JDV85/75 Tradouw Pass. It is characteristically found in the sandstones and can occur in dense masses on steep rock faces.
b.var. longebracteata (Smith) Bayer comb.nov. H. longebracteata Smith, JS.Afr.Bot. 11:75(1945). Scott :127(1985). Type: CAPE-3421(Riversdale): near Stilbaai, Dekenah 18 in Smith 5378 (NBG).
longebracteata: long bracts.
This variety is fairly large and represents the lower-lying larger forms which occur from Bredasdorp across to the Kafferkuils River east of Riversdale. The typical form is from the Stilbaai area where the leaves tend to be suberect with a fairly pronounced end-area. To the west this end-area is less pronounced and both the Bredasdorp and the Swellendam (southwest) forms are reminiscent of the flatter non-retused leaves of H. reticulata.
Distribution: 3420 (Bredasdorp): 5km SW. Swellendam (-AB), Bayer 2420 (NBG), van der Merwe in NBG1184/28, Malherbe in NBG204/41; Napky (-AB) Bruyns in KG37/77 (NBG); 20km N. Bredasdorp (-AC), Venter in NBG868/38, Malherbe in NBG470/42, Smith 3949, 5484 (NBG), Bayer in KG34/70 (NBG); Kransriviermond (-BB), Smith 5752 (NBG); Diepkloof, S. Malgas (-BC), Bayer 2533 (PRE); 3km W. Vermaaklikheid (-BD), Kramer 434 (PRE). 3421(Riversdale): Kafferkuils Bridge (-AB), Smith 6795 (NBG), Bayer 4479 (PRE); S. Riversdale (-AB), Smith 7181, 7204 (NBG); Duiwenhoksriver (-AC), Bayer 2672 (NBG); Brakfontein (-AC), Smith 6107 (NBG), Bayer in KG94/71 (NBG); Botterkloof (-AD), Smith 5378 (BOL,PRE); Kransfontein Farm (-AD), Bohnen 7735 (NBG,PRE); Stilbaai (-AD), Fouche 46 (PRE); Dekenah 18 in Smith 5378 (NBG), Smith 2242, 2811, 5382, 6794 (NBG).
Inadequately located: Heidelberg, Ferguson (BOL).
Haworthia turgida var. longibracteata JDV89/24 north of Stilbaai. Always on rocky steep slopes and in dense clusters.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Very like H. reticulata except for the larger, more turgid and spreading leaves. The flower is much less conspicuous.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Becoming very colourful under stress.
Haworthia turgida var. longibracteata JDV86/6 north of Bredasdorp. A very isolated population.
c.suberecta V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, George district, Mrs Helm in Triebn. 844. Not preserved: Neotype: CAPE-3422(Mossel Bay): Brandwacht (-AA), Bayer in KG631/69 (NBG): H. turgida var. subtuberculata V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, Mossel Bay, Lategan. Not preserved: H. turgida var. pallidifolia Smith, JS.Afr.Bot. 12:10(1946). Type: CAPE-3421(Riversdale): Draaihoek, Albertinia, J. Dekenah 146 in Smith 5714 (NBG).
suberecta: leaves almost erect.
Here the variety is extended to include all these more strongly retused, clump-forming populations which extend all the way from Brandwacht, northeast of Mossel Bay, to Albertinia. This variety is strongly mottled and the leaf ends are slightly truncated and rounded. Smith based his var pallidifolia on a pale-coloured variant which is actually not very common. The Gouritz River form is the most truncated and as, in the typical variety, is highly mottled with semi-translucent dots.
Distribution: 3421 (Riversdale): Draaihoek, Albertinia (-BA), J. Dekenah 146 in Smith 5714 (NBG), Smith 5045, 5490 (NBG), Bayer 4476 (PRE); Weltevrede (-BA), Smith 5537, 7202, 7203 (NBG), Bayer 4471 (NBG); Wydersriver (-BA), Smith 2890 (NBG); Droogerug (-BA), Muir 3006 (BOL); Nuweberg (-BA), Bohnen 9024 (NBG); E. Valsch River Bridge (-BA), Smith 5714, 6912 (NBG), Bayer 4477 (NBG); Gouritz River (-BB), Fouche 48 (PRE), Smith 3857, 3961 (NBG), Bayer 4478 (NBG). 3422 (Mossel Bay): Brandwacht (-AA), Herre in STE6374 (BOL), Bayer in KG631/69 (NBG)
Inadequately located: Mossel Bay, Smith 4952, 5078 (NBG); ex hort Smith 647 (NBG), Naude in NBG1
Haworthia turgida var. suberecta JDV84/74 northwest of Mossel Bay. Leaves abbreviated and rounded at the tips.
Haworthia turgida var. suberecta JDV87/108 Grouritz River. If not so proliferous this would probably be a much more popular plant.
Haworthia turgida var. suberecta JDV94/101 Brandwacht north of Mossel Bay. On stony clay slopes.
Haworthia turgida var. suberecta JDV94/88 northwest of Albertinia. The origin of Smith’s var. pallidifolia in which the leaves are very pale.
38. Haworthia variegata Bolus, J Bot.Soc.S.Afr. :137(1929). V.Poelln., Feddes Repert.Spec.Nov. 27:102(1930). ibid. 44:228(1938). Bayer :165(1976). Bayer :60(1982). Scott :56(1985). Type: Cape, Botterkloof, Mrs. E. Ferguson in Bolus 18900.
variegata: variegated.
Rosette stemless, proliferous, to 4cm φ. Leaves 30-40, erect, slender, lanceolate, margins and keel spined, variegated. Inflorescence simple, slender, to 35cm. Flowers greenish-white with brownish venation, sparse on peduncle.
1982 – This species is for the present only known in the area between Riversdale and Stilbaai. Of the two populations known, one comprises very proliferous plants in rocky situations, and the second sparsely proliferous individuals in sandy areas. The plants have long slender leaves, dark green in colour and strongly variegated. The relationship with other species is obscure and must probably be sought with H. chloracantha. H. variegata and H. divergens are superficially very similar but the latter has incurved leaf tips. Geographically considerations seem to preclude a real relationship between these two species. H. variegata is closely associated with the coastal limestones of the Southwestern Cape. As it is also in the winter rainfall area one can expect that it may present some problems in cultivation.
1999 – Several collections throw more light on this species and the discussion should be considered together with that for H. chloracantha, and H. floribunda. These southern Cape species cannot be considered discrete elements. C. Burgers collected from a very cryptic population on the eastern lower slopes of the Potberg. The plants were very drawn into the gravelly sandstone derived but clayey soil. These were given a manuscript name ‘H. hemicrypta’, applied here at varietal rank. Three other different populations have been recorded all differing markedly from one another and this is expressed in the varieties recognised below. The one at Swellendam is very like H. floribunda var. virescens.
a.var. variegata Known only from Botterkloof and from several kilometres inland in sandy situations. The plants are very variegated. They can only be said to co-occur with one species which is in the same sub-genus, H. turgida.
Distribution: 3421(Riversdale): Droevlakte, Takkiesfontein (-AD), Bohnen 7791 (NBG,PRE); Botterkloof (-AD), Dekenah 19 (PRE), Smith 5384 (NBG), Ferguson in BOL18900; Hoekraal (-AB), Smith 5598 (NBG), Bayer in KG93/71 (NBG); 10km Riversdale to Stilbay (-AD), Minnaar in BOL23214;.
Haworthia variegata var. variegata JDV89/6 Botterkloof. At the type locality the plants form dense clusters.
Haworthia variegata var. variegata JDV87/2 west of Albertinia. Very abundant among rocks in Coastal Fynbos.
Haworthia variegata var. variegata JDV87/2 west of Albertinia. The plants are distinctly variegated
b.var. hemicrypta var.nov. Type: 3420 (Bredasdorp): NE. lower slopes of Potberg (-BC), C. Burgers 2582 (NBG, Holo.).
hemicrypta: half-buried.
Moderately variegated to plain, with leaves long and slender, tending to arch out and then incurved. (A var. variegata foliis longis gracilibus subtiliter variegatis et habitu occultanti differt).
This variety is represented by three populations west of the Breede river with the most inland to the northeast of Swellendam. The plants of the inland populations are quite robust and proliferous. Both the most coastal and the most inland forms are relatively unmarked, whereas the other has distinctive spotting on the lower and outer leaf surfaces.
Distribution: 3420(Bredasdorp): Swellendam (-AB), Tomlinson 13681 (PRE), Ross-Frames in NBG2603/27 (BOL); 3km W. Swellendam (-AB), Smith 3232, 5620 (NBG), DeKok (NBG); Luiperskop (-AD), Venter 1 (NBG); NE. lower slopes of Potberg (-BC), C. Burgers 2582 (NBG), Bayer 2564 (NBG).
Haworthia variegata var. hemicrypta JDV85/10 northeast of Bredasdorp. West of the Breede River are three aberrant populations in which the plants are less conspicuously variegated.
Haworthia variegata var. hemicrypta JDV85/10 northeast of Bredasdorp. This population is just below a Witteberg terricrete cap.
Haworthia variegata var. hemicrypta JDV86/11 northeast of Potberg. In sandstone gravel in low Coastal Fynbos, the plants are extremely cryptic and only the leaf tips are visible.
Leaves broader and shorter and with less conspicuous spination, than the typical variety. (A var. variegata habitu prolificanti et foliis brevioribus pernuerosis aliquantum incurvatis differt).
This variety occurs among fynbos in fairly loose sand amid rocks. It is proliferous and moderately variegated although the fewer leaves are more turgid than expected for this species. There is some evidence of translucence along the margins and at the leaf-tips, particularly in a collection by Venter also from Kathoek. A.R. Mitchell also collected a single specimen of this element from the Potberg Mountain where it was later also noted by P.V. Bruyns. H. heidelbergensis grows in the same vicinity.
Haworthia variegata var. modesta JDV86/68 Kathoek northwest of Bredasdorp. Plants of this original collection do not have the vigour and transclucence of the later collection.
Haworthia variegata var. modesta JDV92/128 Kathoek, northwest of Bredasdorp. Occuring as a much smaller form in the sandstones of the Potberg mountain.
Very proliferous with more leaves which are relatively shorter and incurved than in the typical variety.
The leaves of this variety are rather slender, compared to the more deltoid-shape in the var. modesta. The denser and stronger spination is reminiscent of H. herbacea, but otherwise there is little probability of any close relation with that species.
Haworthia variegata var. petrophila JDV97/24 Karsrivier. Re-collection has indicated that this may have been a spinescent oddity, as the plants were generally much like the var. modesta.
Haworthia variegata var. petrophila JDV86/67 Karsrivier. More spinescent.
This species is named after Jan Vlok, well known naturalist and observer, who has contributed greatly to the knowledge of the flora of the mountains of the Little Karoo and Southern Cape. H. vlokii is a small proliferous species reminiscent of H. maculata from the Langeberg Mountains in the Worcester/Robertson area. The flower tends to be similarly regular with a slightly biarcuate bud. This is also fairly similar to that of H. monticola. The more regular stellate flower, and slightly incurving leaf-tips, are also suggestive of that species which is geographically the most probable affinity. H. vlokii has been recorded at low altitude in Meiringspoort, but this was where it was growing on a huge fallen rock. It is more at home at higher altitude in the mountain pass and was originally also collected on top of the Groot Swartberg Mountains to the east of the Pass. Here it was growing on a north-facing rock wall. There are no co-occurring species although the southern end of Meiringspoort is also home to a depauperate form of H. arachnoidea var. setata. There are several collections, mainly by P.V. Bruyns from the deep valleys on the north side of the Swartbergs. Those to the west of Prince Albert have been included in H. marumiana var. viridis, but one from the eastern Frisgewaagd is a small form of H. vlokii. A lot more exploration and good recording will be required before these mountain-dwellers are better understood. (H. maculata affinis sed foliis parvioribus angunstioribus fuscatioribusque floribus roseo-tinctis, et habitu valde prolificanti differt.)
Rosette acaulescent, 40-50mm diam., proliferous by offset. Stem up to 10mm diam. Leaves spreading to sub-erect, up to 30 X 6-8mm, 3-4mm thick, slightly incurving tips with 2mm long end-awn, surfaces opaque; face convex with 1-2 short rows of inconspicuous opaque whiter dots towards tip; back convex with 3-6 rows of opaque whiter dots, with 1-2 sharper keels bearing spines to 0.6mm long; margins similarly toothed towards apex. Peduncle simple, 1-1.5mm diam., 300-450mm long including raceme, pinkish-brown; base smooth, 3-angled; raceme with 6-18 flowers, 2-4 open; pedicels 4-6mm long, 1mm diam., sterile bracts 5-11, 4-6mm long. Perianth white and brownish-pink, 15mm long, perigon sub-triangular, tepals regular; tube brownish-pink within; buds slightly biarcuate, with rounded tips. Flowering November.
40. Haworthia wittebergensis Barker, JS.Afr.Bot. 8:245(1942). Bayer :168(1976. Bayer :61(1982). Scott :62(1985). Type: CAPE‑3320 (Montagu): Witteberg (‑BA), L. Pieterse in NBG 68214.
wittebergensis: from the Witteberg mountains.
Rosette stemless, slowly proliferous, to 3cm φ. Leaves 20-30, long slender attenuate, amplexicaul at base, grey-green with white spines on margins and keel, coriaceous. Inflorescence simple, slender, to 30cm. Flowers white with green venation, sparsely arranged.
1982 – So many Haworthia species intergrade frustratingly into related species, that it is a relief to find the occasional species which is indisputably discrete. This species has long grey‑green leaves, with a dry papery texture. The leaves are particularly broad at the base where they encircle the stem. H. wittebergensis also avoids direct sunlight and is invariably crammed tightly into rock cracks and crevices. In the dry summers the plants may be very difficult to see because the exposed leaves look no different to small dead tufts of grass. H. wittebergensis is often proliferous but slowly so. It is also a slow grower and requires winter watering. In cultivation the leaves tend to be lighter in colour, and they thicken out and become erect spreading. In the field the leaves may be tightly compacted into a narrow sheaf.
1999 – The distribution seems to be restricted to the mountains to the south of Laingsburg. A relationship with H. pulchella is often proposed when in fact this species and H. blackburniae are probably closer on account of a stem structure which is similar to that of the Hexangulares. The stems have quite woody vascular bundles as opposed to the generally uniformly soft tissue of the stems in the subgenus Haworthia.
Distribution: 3320 (Montagu): Witteberg (‑BC) L. Pieterse in NBG 68214, Smith 5647 (NBG); S. Whitehill (-BC), Compton 15247 (BOL), Lewis & Barker in NBG2251/32 (BOL); SW. Laingsburg (-BC), Bayer 2424 (NBG); Rooinek Pass (-BC), Bruyns in KG13/76 (NBG); Keurfontein (-BD), Bruyns in KG14/75 (NBG).
Inadequately located: Leighton (BOL).
Haworthia wittebergensis JDV91/78 south of Laingsburg. Nearly always in cracks and crevices and with the leaf tips often eaten down.
Haworthia wittebergensis JDV91/78 south of Laingsburg. It does form clumps but is usually restricted by the confinement of the rock cracks.
Haworthia wittebergensis JDV90/67 south of Laingsburg. Difficult to grow because of its winter rain requirement and sensitivity to direct sunlight.
41. Haworthia zantneriana V.Poelln., Cactus J 5:35(1936). V.Poelln., Feddes Repert.Spec.Nov. 41:217(1937). V.Poelln., Desert.Pl.Life 9:90(1937). ibid. Feddes Repert.Spec.Nov. 44:228(1938). Type: Cape, Groot River heights, N. Campherpoort, E. King 111. Not preserved. Lectotype (B&M): icon (B).
zantneriana: in honour of Major Zantner.
Rosette stemless, proliferous, 5-6cm φ. Leaves 20-40, soft, glabrous, attenuate, spreading, pale-green, usually with pellucid white longitudinal marks on the leaves. Inflorescence simple, slender, to 25cm. Flowers white with green venation, sparse.
1982 – H. zantneriana is an unusual species with glabrous smooth leaves marked with pale longitudinal markings. It is proliferous and can form fairly dense clumps. It is usually found on well‑shaded south slopes but only in rock cracks and crevices. The original locality is at Campherspoort near Klipplaat which appears to be at about the mid‑point in the east‑west distribution of the species. There is an indication that the plants get smaller in either direction. Where it occurs north of Willowmore, and also in the Baviaanskloof, there is a strong suggestion that H. zantneriana will be found to be continuous with H. divergens. The plants get smaller with more erect unmarked leaves which are also distinctly and sharply keeled, and have marginal teeth. Towards the east the margins tend to be haired. However, further exploration is needed to determine where the end‑point is in this direction.
1999 – The type of this species is noted to be a collection King 111 from Groot River Heights. There is a collection from further east in the Bolus Herbarium under King 111, but which is from Swanepoelspoort. This specimen is presumed not to be the same as the original and is cited under the var. minor. Not much more has been learned about this species and its affinities. Possibly it continuous with H. gracilis var. viridis in the east and with H. marumiana var. viridis in the west. The more probable affinity is with H. monticola and this is substantiated by two collections. The first is a collection from the Witberg southeast of Willowmore which is nearer to H. monticola in that the leaves are slender, incurved at the tips, and mottled. The texture is relatively soft for that species. The second is from Trompetterspoort, just northeast of Willowmore where the plants really are intermediate between the two species both in appearance and geographically. Note must be taken of the mountain chains along which the respective species are distributed, and along which they may be related.
a.var. zantneriana. Fairly wide spread in the mountains north of the Little Karoo and often quite large, to 8cm tall.
Haworthia zantneriana var. zantneriana JDV97/13 northeast of Willowmore. Quite a robust form with spreading leaves.
Haworthia zantneriana var. zantneriana JDV93/42 north of Campherpoort. The normal larger form often has white keels and margins, and some similar flecking on the leaf surfaces.
b.var. minor var.nov. Type: CAPE-3323 (Willowmore): near Miller Station (-BB), Bayer 1702 (NBG, Holo.).
minor: small.
Small, to 5cm, without markings on the leaves. (A var. zantneriana foliis parvioribus immaculatis differt).
The specimen Bruyns 7071 cited below from near Willowmore (Trompetterspoort) is intermediate between this species and H. monticola as mentioned earlier.
Distribution: 3323 (Willowmore): N. Redcliffe (-BA), Bruyns 7061 (BOL); Trompetterspoort (-BA), Bruyns 7071 (BOL); near Miller Station (-BB), Bayer 1702 (NBG); Bakenskop (-BB), Mitchell 354 (NBG). Swanepoelspoort (-BB), King 111 (BOL).
Haworthia zantneriana var. minor JDV85/26 Miller Station. A smaller form with less conspicuous markings.
Haworthia zantneriana var. minor JDV97/17 northeast of Willowmore. A small form transitional to H. monticola.
42. Haworthia attenuata Haw., Syn.Pl.Succ. :92(1812). Haw., Rev.Pl.Succ. :54(1821). Berger, Das Planzen. :93(1908). Bayer :100(1976). Bayer :63(1982). Scott :15(1985). Aloe attenuata Haw., Trans.Linn.Soc. 7:11(1804). Salm-Dyck, Monogr. 6:t12(1834). Aloe radula sensu Ker-G., Curtis’ Bot. Mag. t.1345(1811). Type: Not preserved. Neotype (designated here): icon t1345, Curtis’ Bot. Mag.: H. clariperla Haw., Phil.Mag. :186(1928). Aloe attenuata var. clariperla (Haw) Salm-Dyck, Monogr. 6:12b (1834). H. attenuata var. clariperla (Haw.) Baker, J Linn.Soc. :204(1880). Farden, Cactus J 8:34(1939): H. attenuata fa clariperla (Haw.) Bayer :107(1976). Bayer :63(1982): Type: Not preserved. Neotype (designated here): icon, 6:12b, Salm-Dyck, Monogr.: H. fasciata var. caespitosa Berger, Das Pflanzen. 33:92(1908). Type: Not preserved: H. brittenianae V.Poelln., Feddes Repert.Spec.Nov. 41:196(1937). H. attenuata var. britteniana V.Poelln., Kakt.u.and.Sukk. 10:166(1937). H. attenuata fa britteniana (V.Poelln.) Bayer :104(1976). Bayer :63(1982). Type: Cape, Oudtshoorn, Mrs van der Bijl in Triebn. 807. Not preserved. Neotype (B&M)): icon (B). Epitype: Plutosvale, Smith 1118 (NBG). H. attenuata var. odonoghueana Farden, Cactus J 8:34(1939), et vars. linearis, uitewaaliana, deltoidea, minissima, inusitata. Types: not preserved.
attenuata: tapering leaves.
Rosette stemless, proliferous, to 130mm tall. Leaves to 130 X 15mm, attenuate, spreading, lanceolate-deltoid, surfaces scabrid with distinct raised non-confluent tubercles. Inflorescence sparsely branched, lax. Flowers tepals fused, tube straight, lower inner tepals revolute.
1982 -This species is widespread in the Eastern Cape from beyond the Fish River in the east, to the Humansdorp area in the west. Old records suggest that it was collected near Umtata in the Transkei and also in the Kei River valley. It may appear to be closely related to H. fasciata but it is distinguished by having longer, thinner and generally more spreading leaves. The leaf faces are also always tubercled. H. attenuata occurs in the drier Valley Bushveld vegetation. In the Hankey area, H. attenuata and H. radula grow together. There do not appear to be any direct and obvious intermediates between very similar species but breeding experiments may prove them to be the same. H. attenuata fa clariperla is very common in gardens and collections throughout South Africa and is recognised by the large white confluent tubercles in bands across the back of the leaves. The original origin of this form is not known but it has been collected at Fort Brown. H. attenuata fa britteniae was collected in Plutosvale and is recognised by its many large white discrete tubercles. Farden recognised a host of varieties based on minor differences. Smith (1950) pointed out some of the difficulties which arise when one goes to this extreme, saying that it would be possible, on the basis of leaf characters, to derive several varieties from the same plant.
1999 – The recognition of two separate forms is abandoned, and it is suggested that variants are referred to by locality of origin. There is as yet no further confirmation of a distribution extending to beyond the Fish River in the east. Breuer and Metzing neotypify the species name by a specimen which is cited under the var. radula in this work. There really is no need to depart from early illustrations for typification purposes for which there are several precedents. The species is well known, and Breuer and Metzing have introduced the first confusion of identity in the history of the species.
a.var. attenuata. This variety is recognised from necessity arising from the inclusion of H. radula within this species, and in preference to recognising the many other varieties already in the history of the species.
Distribution: 3226 (Fort Beaufort): Middeldrift (-DD), Smith 3511 (NBG). 3324 (Steytlerville); E. Hankey (-DD), Smith 2910 (NBG); Kleinriver (-DD), Smith 7380 (NBG). 3325(Port Elizabeth): Enon (-BC), Bruyns in KG40/77 (NBG); 21km Port Elizabeth to Uitenhage (-CD), Smith 5820 (NBG); Addo (-DA), Barker 5090 (NBG), Smith 3549 (NBG); (-DC), Britten in PRE 34850; Swartkops (-DC), (BOL). 3326 (Grahamstown): Plutosvale (-BA), Marloth 5962 (PRE), Harcourt Wood in BOL20729, Britten in PRE 34891, NBG338/34, Smith 914, 1118 (NBG); Fort Brown (-BA), Smith 3512, 3825 (NBG); Glenelg (-BB), Smith 5406 (NBG); Ballinafad (-BB), Smith 5400, 5407 (NBG); E. Fish River (-BB), Smith 5409, 5410 (NBG).
Inadequately located: Near Grahamstown, Luyt in NBG304/45 (NBG); Albany dist, Britten in PRE 34892; ex hort, Simpson in NBG233/42, Peers in NBG34/36, Howard in NBG82/25, Meterlerkamp in NBG1450/29 (NBG), van der Merwe in NBG1696/29 (NBG), Marloth 4210, 5963 (PRE); Kingwilliamstown, Carter (BOL); Sandile Koppie, Grant & Blenkins in NBG2257/26 (BOL); Smith 617, 2822, 3225, Bayer 169 (NBG), Henderson 1399 (NBG).
Haworthia attenuata var. attenuata JDV90/42 Addo. The tubercles are conspicuously white on both leaf surfaces. The leaves have no fibres although viscid strands can be seen on breaking the leaves.
Haworthia attenuata var. attenuata MBB6831 Hankey. Not surprisingly the tubercles can be smaller and evenly spaced.
b. var. radula (Jacq.) Bayer comb.nov. Haworthia radula (Jacq.) Haw., Syn.Pl.Succ. :8(1812). Berger, Das Pflanzen. 38:94(1908). Bayer :148(1976). Bayer :71(1982). Scott :19(1985). Aloe radula Jacq., Hort.Schoenb. 4:11.t422(1804). Salm-Dyck, Monogr. 6:t8(1834). Type: icon, t422, Jacq., Hort.Schoenb. Epitype (B&M): CAPE-3324 (Steytlerville): 1.5km E. Hankey (-DD), Smith 3190 (NBG).
radula: like a rasp.
1982 – H. radula is only known from the immediate vicinity of Hankey in the Humansdorp area where it grows together with H. attenuata. There does not appear to be a random association, however, and the two species appear to be discrete in the small localised population as far as is known. H. radula always has many minute crowded white tubercles in contrast to the larger fewer tubercles on the leaves of H. attenuata. Despite the variability of the latter species there do not seem to be obvious intermediates between the two. It is a very attractive plant in cultivation as the many tubercles give the plant a silvery appearance.
1999 – On reflection, some of the above comments are not strictly true. There is some difficulty in deciding if a plant properly qualifies as ‘radula’ or not. There is no evidence at all of range and distribution for the element outside of H. attenuata; perhaps even varietal rank is unnecessary.
Distribution: 3324 (Steytlerville): Sandland, 20 km E. of Patensie (‑DA), P.L. Perry 660 (NBG); Humansdorp (-DD), Marloth 5961 (PRE); Hankey (‑DD), Smith 3190, 7382 (NBG), Holland 4141 (PRE); 1.5km E. Hankey (-DD), Smith 2933, 3190 (NBG); 1.5km N. Hankey (-DD), Smith 3692 (NBG); 1.5km SE. Patensie (-DD), Smith 3182 (NBG).
Inadequately located: Cape, Marloth 10086 (PRE); ex hort, Buckland in NBG428/31 (BOL, NBG), Malherbe in NBG486/42 (NBG); Hansmeykraal, Fourcade 3550 (BOL).
Haworthia attenuata var. radula MBB6803. The tubercles are small and plentiful and the leaves thus appear scabrid rather than tuberculate.
Rosette stemless, solitary, to 6cm φ. Leaves 5-11, with flattened, retused end-area, opaque and slightly scabrid with small raised tubercles above. Inflorescence simple, slender, florets sparse. Flowers slender, tepals fused and with revolute tips.
1982 – H. bruynsii is the most remarkable discovery since H. truncata in 1910. The general appearance of the plants and their locality immediately suggest H. springbokvlakensis. However, the surface of the leaf-end areas is more scabrid and often more acutely margined as in some forms of H. emelyae. The surprise comes in the inflorescence as the peduncle is long and wiry and the florets are singularly characteristic of the subgenus Hexangulares, i.e. six‑sided at the base, exposed mid‑ribs of inner perianth lobes, straight style and florets slowly spiralled on the peduncle. Had only one plant been seen it may have been considered a hybrid of some kind. However, H. bruynsii occurs at two distinct localities as is so far known, and there is no evidence of hybridisation. The plants have only 5‑11 leaves when full grown, and they are sunken in the ground under small bushes in the same way as H. springbokvlakensis and also occurring with Lithops localis as in this species.
1999 – This species is now well-known southeast of Steytlerville and speculation about its relationships continues. J.D. Venter has suggested that the affinity is with H. sordida. This is plausible as that species does show signs of abbreviated leaves and the tendency towards a truncated end-area, in collections east of H. bruynsii localities. H. sordida does, however, also occur in its normal large and erect form nearby at Springbokvlakte and also at Kleinpoort a little to the north. The comparison with H. emelyae above should now read H. bayeri.
44. Haworthia coarctata Haw., Phil.Mag. 44:301(1824). Baker, JLinn.Soc.Bot. 18:202(1880). Salm Dyck, Monogr. 6:f17(1936). V.Poelln., Feddes Rep.Spec.Nov. 43:108(1938). Bayer, Natn.Cact.Succ.J 28:80(1973): Bayer :107(1976). Bayer :64(1982). Scott :48(1985). Type: Not preserved. Neotype (designated here): icon (K). Epitype (ex B&M): Grahamstown to Bathurst, Smith 7092 (NBG). H. chalwinii Marl.et Berg., Notizbl.Bot.Gart.Mus.Berl. 4:247(1906). Type: Cape, Graaff Reinet, Marloth 4051 (K): H. reinwardtii var. conspicua V.Poelln., Feddes Repert.Spec.Nov. 41:210(1937). Type: Cape, Port Elizabeth to Alexandria road, Archibald 85 in Long 346. Not preserved: H. reinwardtii var. fallax idem :209(1937). H. fallax idem. 31:83(1932). Type: Cape, Grahamstown, STE6633. Not preserved: H. reinwardtii var. pseudocoarctata V.Poelln., Beitr.Sukk. 2:43(1940). H. coarctata var. haworthii fa pseudocoarctata (V.Poelln.) Res., Mems.Soc.Broteriana: Succ.Afr. 3:84(1943). H. coarctata var. haworthii Res. ibid.: H. coarctata var. kraussii ibid.: H. reinwardtii var. committeesensis Smith, JS.Afr.Bot 9:93(1943). Type: Cape, Albany Div. Smith 551 (NBG): H. reinwardtii var. huntsdriftensis ibid. 10:14(1944). Type: Cape, Albany Div., Smith 3849 (NBG): H. fulva idem. 9:101(1943). Type: Cape, Bathurst Dist., Smith 3380 (NBG): H. musculina ibid. 14:49(1948). Type: Cape, Bathurst Dist., Smith 5118 (NBG): H. greenii var. silvicola ibid. 9:103(1943). Type: Cape, Bathurst Dist., Smith 3378 (NBG).
coarctata: leaves pressed together.
Rosette to 120mm φ, caulescent, proliferating. Leaves many, to 70 X 20mm, ratio stem diameter to leaf width 1:1.7, erect spreading or incurved, scabrid, brownish-green, usually with rounded tubercles. Inflorescence simple or occasionally compound, to 300mm. Flowers tepals fused, tube straight, lower inner tepals revolute.
1982 – This is one of the stem‑forming species occurring west of the Fish River north of Grahamstown and extending to near Port Elizabeth. It has very often been confused with H. reinwardtii as the above synonymy suggests. However, there is a distinction based on both geographical distribution and morphological discontinuity. This is discussed in detail in Bayer (1973). Essentially there is a difference between the two species reflected in the number of leaves per unit length of stem. In H. coarctata the ratio of stem diameter to leaf width is 1:1.7 while in H. reinwardtii leaf width more nearly equals stem diameter and the ratio is 1:1.2. In H. coarctata the leaves are less densely arranged on the stem and also thicker. Another difference is that in H. coarctata the tubercles are smaller and more smoothly rounded, whereas in H. reinwardtii they are frequently large and tend to be flattened and whiter. H. coarctata has many striking forms and, possibly because of vegetative reproduction, populations tend to be very uniform. The subspecies coarctata contains the var. greenii, which is a glabrous variety occurring only at Howiesonspoort. The var. tenuis is a very slender variation with grossly elongated stems occurring near Alexandria in the south. There are countless other forms such as that which may have been described as H. chalwinii from along the Kowie River; the beautifully silvery tubercled forms from the Salem area and the very large hexaploids from the western and northern populations. The subsp. adelaidensis is a smaller element which occurs in the False Macchia around Grahamstown, perhaps the counterpart of H. reinwardtii var. brevicula towards the Fish River. The var. bellula must have been an aberrant clone because no such really small forms were found on a visit to the type locality. The western limit of H. coarctata is not known but it appears to be before the 26th line of longitude, east of Paterson, where the plants are still large and probably hexaploid. Odd and unconfirmed collections from between here and Uitenhage, notably at Addo, suggest that depauperate forms may be found in that area.
1999 – The changes here are the abandonment of the sub-species in favour of varietal rank. Possibly the correct solution is actually the inclusion of H. coarctata and H. reinwardtii in a single species. It is sometimes difficult to separate the two elements and there is an indication of their single identity in the Port Alfred to Alexandria vicinity. There may also be a similar problem in the area above and to the west of Hunt’s Drift. This is where the var. huntsdriftensis was collected and I was unable to find anything other than H. reinwardtii var. brevicula in that general area. While many species and varieties have been named within this group it is obvious from those and from new collections, that an endless array of different forms could be selected. This is in fact what G.G. Smith was in the process of doing as testified by the species fulva and musculina and greenii var. silvicola from the one farm, Hopewell. Smith also identified plants from this farm as H. chalwinii and the typical H. coarctata. A similar situation occurred with the Kaffirdrift populations of H. reinwardtii. The species citations show that Smith collected and numbered individual clones. During the years between collection and the making of the herbarium specimen, these plants were observed and described in minute detail. It is obvious that Smith was in the process of identifying new species and varieties from this array of material. H. chalwinii was described from a plant sent by Marloth from Graaff-Reinet. This is a form of H. coarctata which has a much narrower rosette with shorter stubby leaves. I have collected similar forms along the Kowie River, and Smith’s identification for Hopewell is not the only evidence suggesting occurrence in other populations. The most interesting new record is that by Mr Massyn in the Motherwell area of Port Elizabeth. This record is of a large form refuting the earlier suggestion of depauperate forms to the west. The distribution of the species is still complementary to that of H. fasciata, however, and fuller distribution records of both species are still necessary.
Dr D. Cutler (priv.comm.) said that he did not support he differences between H. coarctata and H. reinwardtii as set out by me in 1973. However, there is no evidence against my suggestion even though Col. Scott also disregards that arrangement altogether. If these two are correct, then the rational alternative is that only one species is involved. This is a solution which I would accept with the proviso that the geographical subdivision of that species is possible. The vars coarctata and adelaidensis are in two cases, viz. Hellspoort and Gowies Farm, reported from the same locality, although the latter identification is from herbarium specimens only. H. baccata is a name given by Smith to a plant ostensibly from southwest of Stutterheim (Smith 3572, wrongly given as 3782 by myself in 1982, and as 3527 by Scott 1985). The NBG herbarium specimen is annotated by McClaren as an apparent introduction to the area. It was never seen there by Smith and on visiting the given site, I could neither find it nor concede that the species could grow there. I noted in 1982 that a specimen noted by me as Smith 3782, was also in the Berlin Dahlem herbarium, which did not accord with the NBG specimen. Confusion is very easily sown.
a.var. coarctata This is the main body of the species west of the Fish River. Three taxonomic variants are recognised as is evident below. Of these, the fa greenii is very localised. The two varieties have more substance, but the typical variety still encompasses a substantial range of variation. Again I see no reason to depart from the stable historical record of the species to find a neotype.
Distribution: 3325 (Port Elizabeth): Motherwell (-DA), Massyn (NBG); Addo (-DA), Fourcade 39 (NBG), Smith 2774 (NBG); Orlando (-DB), Smith 5348, Bayer 1371 (NBG). 3326 (Grahamstown): Hellspoort (-AB), Bayer 1352 (NBG); Piggot Bridge road (-AB), Britten in PRE 34837; Bushmans River Poort, Paynes farm (-AC), Archibald 5595 (PRE); Rockdale, Highlands (-AC), Dyer 2213 (BOL); Salem to Alexandria (-AD), Smith 3573, 3955, 4010 (NBG), Stayner (NBG); NE. Salem (-AD), Smith 3412, 3414, 3415 (NBG); Yellowwoods (-AD), Bayer 1372 (NBG); Howiesonspoort (-AD), Dyer 2507, 2508 (BOL), Reynolds in NBG987/34 (NBG); Rooidrift (-BA), Bayer 1367 (NBG); Fort Brown (-BA), Walton in NBG141/24 (BOL), Smith 5825 (NBG), Walton in NBG141/24; Gowie’s Farm (-BA), Smith 5625a (NBG); E. Plutosvale (-BA), Bayer 1368 (NBG); Fletcher’s Farm (-BA), Smith 5423, 5424, 5425 (NBG); E. Committees (-BB), Smith 7421 (NBG); W. Committees (-BB), Smith 551 (NBG, PRE); W. Breakfast Vlei (-BB), Smith 3959 (NBG); Brooklands (-BC), Bayer 1357 (NBG); Manley Flats (-BC), Fourcade 195, Bayer 1358 (NBG); Vaalvlei (-BC), Bayer 1360 (NBG); Hills SE. Grahamstown (-BC), Britten in PRE 34834; 11km Grahamstown to East London (-BC), Wells 2661 (PRE); SE. Grahamstown (-BC), Smith 7092, 7092a,b,c,d,f,h (NBG); W. Hunt’s Drift (-BD), Smith 3849 (NBG ,PRE), Smith 6818, 6818a,b (NBG); Bathurst (‑BD), Smith 7092 (NBG); Woodbury (CA), Bayer 1370 (NBG); Salem to Alexandria (-CB), Smith 3413, 3410, 3411 (NBG), Bayer 1363 (NBG); 24km W. Port Alfred (-DA), Smith 3378 (PRE); Kowie (-DA), Bayer 1359 (NBG); S. Vaalvlei (-DA), Bayer 1361 (NBG); Hopewell, 20km W. Port Alfred (-DA), Fourcade 119 (NBG), Smith 3380 (NBG, PRE), Smith 3378, 5114, 5115, 5116, 5117, 5118 5119 5120 (NBG), Bayer 1364, 1366; 24km WNW. Port Alfred (-DA), Smith 5115 (PRE); Near Ghio Bridge (-DA), Smith 7342 (NBG); Hopewell (-DA), Acocks 11054 (PRE); Glennismoyle (-DB), Smith 3416, 7491 (NBG) Fairfax (-DB), Bayer 1194 (NBG). 3327 (East London): E. Hunt’s Drift (-AC), Smith 5667 (NBG).
Inadequately located: Albany, Blackbeard in NBG 582/25; Brakfontein, Alexandria, Galpin (BOL); Alexandria, Archibald in NBG1322/37 (NBG); ex hort Marloth 4215 (PRE), Repton 61 (PRE), Whitehill (NBG).
Haworthia coarctata var. coarctata JDV90/46 Howiesonspoort. The density of leaf arrangement and tubercle pattern are a little different in each population.
Haworthia coarctata var. coarctata JDV97/76 Belmont, east of Grahamstown. Often in huge mats with individual stems to 30cm.
b. fa greenii (Baker) Bayer comb.nov. H. greenii ssp. coarctata var. greenii (Baker) Bayer, Natn.Cact.Succ.J 28:80(1973). Bayer :120(1976). Bayer :65(1982). H. greenii Baker, JLinn.Soc. 18:202(1880). Scott :50(1985). Type: Cape, Cooper 1860 (K): H. peacockii Baker, JLinn.Soc. 18:202(1880). Type: ex hort. Kew. Not preserved: H. greenii fa bakeri Res., Mems.Soc.Broteriana: Succ.Afr. 3:87(1943). ex hort. Lisbon. Not preserved: H. greenii fa minor ibid. Type: ex hort. Lisbon. Not preserved.
greenii: for C.G. or G.H. Green.
This variant is only known from Howiesonspoort, and the normal tubercled variety is also present both to the east and west of the specific site of this glabrous variant. Col. Scott suggests a wider distribution but there is absolutely nothing in the herbarium record to support any notion that this is an independent entity. Even the criteria (no tubercles) is really inconsequential in a genus where many species may have tubercled or non-tubercled variants. The var. silvicola is obviously also an insignificant variant from the farm Hopewell where Smith collected many variants which he translated into species and varieties.
Distribution: 3326 (Grahamstown): Howiesonspoort (‑AD), Howiesonspoort, Dyer 2506, Reynolds in NBG 68038, Reynolds in NBG987/34, (BOL, NBG), Dyer 2507, 2508 (NBG), Smith 3178, 3524, 3525 5303 (NBG).
Inadequately located: ex hort. Whitehill, Smith 7448 (NBG).
Haworthia coarctata var. coarctata fa. greenii JDV90/46 Howiesonspoort. This form has no tubercles and its distribution is limited to a small area west of Grahamstown.
c. var. adelaidensis (V.Poelln.) Bayer comb.nov. H. coarctata ssp. adelaidensis (V.Poelln.) Bayer, Natn.Cact.Succ.J 28:86(1973). Bayer :94(1976). Bayer :65(1982). H. reinwardtii var. adelaidensis V.Poelln., Beitr.Sukk. 2:43(1940). Type: Cape, Adelaide, Armstrong. Not preserved. Neotype (B&M): icon (B): H. reinwardtii var. riebeeckensis Smith, JS.Afr.Bot. 10:16(1944). Type: Cape, Albany Div., Smith 5218 (NBG): H. reinwardtii var. bellula ibid. 11:70(1945). Scott, Nat.Cact.Succ.J 36:37(1981). Scott :47(1985). Type: Cape, Albany Div., Smith 3137 (NBG).
adelaidensis: from Adelaide, Cape.
As noted above there are two records which indicate that the typical variety, and this one, occur in close proximity. This may also be true for a site to the southeast of Grahamstown. The variant bellula is obviously a single clone which was collected my Miss Britten. A second clone was annotated ‘long-leaves’. Smith himself subsequently appears to have visited the site, or at least the near vicinity, twice. He does not record collecting a similar plant, nor do his specimens show this. Both myself and G. Marx have failed to find a similar clone at the given site. There is very little difference between this variety and var. tenuis except that the latter does develop much longer stems with age.
Distribution: 3326 (Grahamstown): 8km W. Riebeek East (-AA), Smith 7399 (NBG); Willowfountain (-AA), Bayer 1355 (NBG); Near Riebeek East (-AA), Smith 5218 (BOL, NBG, PRE); Brakkloof (-AB), Marx 43 (NBG); Hellspoort (-AB), Smith 355 (NBG, PRE), Bayer 1351 (NBG); Top Plutosvale (-BA), Smith 5426, 5427, 5428, 5429, 5430 (NBG); Glendew (-BA), Smith 5421 (NBG); Gowie’s Farm (-BA), Smith 5625 (NBG); Striata Hill (-BA), Smith 2431 (NBG); Bothas Hill (-BA), Barnes (BOL); Near top Queens Road (BA), Smith 2839 (NBG, PRE); Commonage (-BC), Smith 7448 (NBG); Queens Road (‑BC), Smith 2839 (NBG), Smith 7427 (NBG); NW. Grahamstown (-BC), Smith 3137 (BOL, NBG), Smith 3137a, 5626, 5626a (NBG), Bayer 1354 (NBG); Ainsley’s Farm, Trappes Valley (-BD), Smith 6782 (NBG).
Inadequately located: Albany, Luyt in NBG307/45 (NBG); Henderson 1450 (NBG), Rennie in NBG 1268/32.
Haworthia coarctata var. adelaidensis JDV94/53 northwest of Grahamstown. The small form named as bellula.
Haworthia coarctata var. adelaidensis JDV97/65 northeast of Grahamstown. The plants do vary in size but stem-length is usually restricted.
d. var. tenuis (Smith) Bayer comb.nov. H. coarctata ssp. coarctata var. tenuis (Smith) Bayer, Natn.Cact.Succ.J 28:80(1973). Bayer :106(1976). Bayer :65(1982). H. reinwardtii var. tenuis Smith, JS.Afr.Bot. 14:51(1948). Type: Cape, Alexandria Dist. Smith 3420 (NBG).
tenuis: slender.
This variety is generally about half the diameter of the more typical variety and the leaves are more slender – which would separate it from the fa chalwinii. There do appear to be other populations of such slender leaved forms along the lower Bushman’s River, which should be distinguishable from H. reinwardtii by both tubercle character and leaf density on the stem.
Distribution: 3326 (Grahamstown): Harvestvale, Alexandria (‑DA), Smith 3417, 3418, 3419, 3420 (NBG), Holland 3817 (BOL).
Inadequately located: Bushmans River, Britten (NBG).
Haworthia coarctata var. tenuis. This relatively delicate variant is prone to die back at the leaf tips unless it is evenly watered or well shaded.
Haworthia coarctata var. tenuis PVB Harvestvale. Mat forming with long slender stems which are very similar to var. adelaidensis, but grow very much longer.
45. Haworthia fasciata (Willd.) Haw., Suppl.Pl.Succ. :57(1819) in fort. H. fasciata var. major Haw., Syn.Pl.Succ. :58(1819) Haw., Revis. :54(1821). Baker, JLinn.Soc.Bot. 18:204(1880). Bayer :116(1976). Bayer :66(1982). Scott :20(1985). Apicra fasciata Willd., Ges.Naturf.Fr.Berl.Mag. S:270(1811). Aloe fasciata (Willd.) Salm Dyck, Monogr. 6:f15(1837). Type: None known. Neotype (designated here): icon, f15, Salm Dyck 6(1837). Epitype (ex B&M): Hankey, Stayner (NBG): Aloe fasciata var. major Salm Dyck, Monogr. 6:fl5B(1837). H. fasciata var. major (S.D.) V.Poelln., Feddes Repert.Spec.Nov. 43:95(1938). Type: icon, Salm Dyck 6:f15B(1837): H. fasciata var. subconfluens V.Poelln., Kakteenk. 9:133(1937). H. fasciata fa subconfluens V.Poelln., Feddes Repert.Spec.Nov. 43:95(1938). Type: Cape, Humansdorp, Mrs I. King. Not preserved: H. fasciata fa ovatolanceolata idem 43:96(1938). Type: Cape, Humansdorp, Mrs I. King. Not preserved: H. fasciata fa sparsa ibid. 43:96(1938). Type: Cape, Witteklip near Uitenhage, Long 479. Not preserved: H. fasciata fa variabilis ibid. 43:96(1938). Type: Cape, Elandsriver road, Cook in Long 443. Not preserved: H. fasciata fa vanstaadenensis ibid. 43:97(1938). Type: Cape, Vanstaadens Pass, Long 443. Not preserved: H. browniana V.Poelln., Desert Pl.Life 9:102(1937). V.Poelln., Feddes Repert.Spec.Nov. 44.212(1938). H. fasciata fa browniana (V.Poelln.) Bayer :105(1976). Bayer :66(1982). Scott :21(1985). Type: Cape, Uitenhage, Armstrong. Not preserved. Lectotype (B&M): icon (B).
fasciata: barred.
Rosette to 150mm φ, stemless, proliferating. Leaves 60-80, erect, to 60 X 15mm, incurved, scabrid, white tubercles on underside only. Inflorescence simple or occasionally compound, to 300mm. Flowers tepals fused, tube straight, lower inner tepals revolute.
1982 – H. fasciata occurs in False Sclerophyllous Shrub vegetation around Port Elizabeth and east of the Gamtoos river valley. Unlike H. attenuata and H. radula which have lanceolate leaves with tuberculate leaf faces, in H. fasciata the leaves are more deltoid and the leaf faces smooth. The leaves also tend to curve inwards so that the plants can resemble young stemless rosettes of H. coarctata or H. reinwardtii. The fa browniana occurs as only one small population north of Uitenhage which has been reported to be a vegetatively propagated clone (an interchange heterozygote; Brandham, 1981). This form tends to be caulescent, brownish in colour, and with longer and broader leaves than is usual for the species. H. fasciata is more difficult in cultivation than H. attenuata and this may be because it is far less proliferous and because it occurs in acid soils.
1999 – There is a curious twist in the synonomy of this species because Haworth seemed to have actually been unaware of Wiildenow’s description and applied the same epithet to a plant fortuitously the same. It appears that his species was actually H. reinwardtii and his variety major was synonymous with Willdenow’s species. Somebody else can resolve the synonymy in a better way if they choose. As is also obvious from the synonymy, there is also quite a considerable variation in this species although its distribution is relatively closely confined to the Port Elizabeth area. Plants from east of Uitenhage can be quite heavily tubercled and are more attractive in cultivation than most members of the species. Dr. Hayashi (priv. comm.) refutes the contention that the forma browniana comprises a single clone. Here it is no longer formally upheld as a taxon although it is an obvious variant, because recognition of variants of this kind in a consistent way would result in an excessive proliferation of names. Although the normal distinction from H. attenuata is on the basis of a tubercle-free upper leaf surface, H. fasciata has fibres in the leaf which must constitute a more profound difference between the two species. Fibres are also present in the leaves of H. longiana, H. coarctata and H. glauca and this character may suggest a real division in the subgenus.
Distribution: 3324 (Steytlerville): Moordenaarskloof (-CC), Stayner (NBG); W. Assegaaibos (-CC). Smith 896 (NBG); Witteklip (-CD), Fourcade 124 (NBG); Near Assegaaibos (-CD), Smith 2071 (NBG); Assegaaibos (-CD), Fourcade 2409 (BOL); E. Patensie (-DD), Smith 2907 (NBG); Gamtoos Bridge (‑DD), Stayner in NBG68720, Fourcade 10, 11 (NBG), Smith 5799, 7265a (NBG); Hankey (DD), Fourcade 9 (NBG), Smith 2295 (NBG), Smith 5745 (NBG), Stayner in KG 352/62;; N. Hankey (-DD), Smith 2929, 3693 (NBG); E. Hankey (-DD), Smith 3180, 5797, 7067 (NBG), Stayner (NBG); Hankey to Humansdorp (-DD), Smith 6812 (NBG); W. Gamtoos Bridge (-DD), Smith 5795 (NBG); NE. Zuurbron (-DD), Smith 3670 (NBG), Bayer & Bruyns 6552 (NBG). 3325(Port Elizabeth): Spring Range (-CB), Bayer 886 (NBG); Hillwacht (-CB), Bayer in KG155/73 (NBG); Uitenhage (‑CB), Stayner in NBG68919; Loerie (-CC), Fourcade 8 (NBG), Stayner (NBG); Vanstaadens Pass (-CC), Fourcade 125 (NBG), Smith 5796, 6133 (NBG); 8km W. Thornhill (-CC), Smith 2866, 3179, 3183, 3183a; 8km E. Hankey (-CC), Bayer 887 (NBG); Bulkrivier (-CC), Fourcade 123 (NBG); Thornhill (-CC), Coates (BOL); Despatch (-CD), Marloth 13166 (PRE); W. Uitenhage (-CD), Smith 706 (PRE); Korsten Hill (-DC), Smith 5798 (NBG); Port Elizabeth (-DC), Stayner (NBG); Cotswold (-DC), Branch 365 (NBG); W. Bethelsdorp (-DC), Smith 5821, 5822 (NBG); Baakens Valley (-DC), Cruden 354 (PRE); Korsten Hill (-DC), G.F. Smith 219 (PRE).
Inadequately located: Steytlerville, Compton in NBG1043/47; Nelspoort, Pearson 520; Port Elizabeth, Grey (BOL); Humansdorp, Coates (BOL); Lavis in NBG2694/27 (BOL), Smith 3241 (NBG), Stayner (NBG), Fourcade 105 (NBG).
Haworthia fasciata JDV90/56 Gamtoos Bridge. A rather large form like the variety described from Vanstadens Pass viz. “ovato-laceolata”.
Haworthia fasciata JDV94/117 east of Hankey. The upper leaf surfaces are normally without tubercles. The leaves are fibrous.
Haworthia fasciata JDV90/41 Bethelsdorp. Usually a fynbos species, many habitats in and around Port Elizabeth hav been destroyed by urban sprawl.
Haworthia fasciata JDV96/58 Zuurbron. The plants here are slightly caulescent and the tubercles may form transverse bars.
Haworthia fasciata JDV90/39 north of Uitenhage. This is the robust “forma browniana”.
Rosette stemless, proliferous, to 120mm tall. Leaves to 80 X 15mm, attenuate, spreading, lanceolate-deltoid, scabrid with or without distinct raised non-confluent excrescences. Inflorescence sparsely branched, lax. Flowers tepals fused, tube straight, lower inner tepals revolute.
1982 – Strangely enough this is one of the most common haworthias found in collections despite never having been seen or collected in the wild. The assumption has always been that it is a garden hybrid but Brandham (unpublished) maintains, on the basis of cytological examination, that this is not so. H. glabrata is not actually smooth. The small tubercles are concolorous and the plants are a brightish green colour. The leaves are lanceolate as in H. attenuata but they are much rounder in cross‑section, larger at the base, and fleshier in texture. It is very proliferous and easy to grow.
1999 – This species was discovered in the wild by P.V. Bruyns in the same general area that Aloe reynoldsii grows, and together with H. cymbiformis var. setulifera. Thus Dr Brandhams evidence and conclusions are correct. Dr Gideon D. Smith was fascinated by this species and wrote (1989) quite extensively about it. There is only one other collection which could be this species, and that is the one cited below from between the Kei and Tsomo Rivers. There is also a collection in the Bolus Herbarium from ‘Thabase near Umtata’. This consists of a single leaf which suggests Hexangulares rather than Haworthia.
Inadequately located: Between Kei and Tsomo Rivers, Holmes in BOL71340; ex hort Grenfell in NBG871/35, Smith 2004, 2770, 4943, 4944 (NBG), Bayer 170 (NBG).
Haworthia glabrata PVB4400 Bashee River. Only recently re-collected. The leaves are thicker and fleshier at the base than in H. attenuata.
Haworthia glabrata PVB4400 Bashee River. The older forms in cultivation may have had more erect leaves.
47. Haworthia glauca Baker, JLinn. Soc. 18:203(1880). V.Poelln., Cact.Succ.J(U.S.) 5:30(1936). ibid., Feddes Repert.Spec.Nov. 43:108(1938). Bayer :118(1976). Bayer :67(1982). Scott :49(1985). Type: Cape, Zuurberg Pass, Cooper (K): H. carrissoi Res., Bolm.Soc.Broteriana 15:161(1941). Type: Cape, Zuurberg Pass. Not preserved.
glauca: glaucous, powdery, bluish-green.
Rosette to 80mm φ, caulescent, proliferating. Leaves many, to 60 X 15mm, erect spreading or incurved, scabrid, glaucous grey-green, with or without tubercles. Inflorescence simple or occasionally compound, to 300mm. Flowers tepals fused, tube straight. Lower inner tepals revolute.
1982 – Baker gave the locality for H. glauca as ‘Orange Free State’ which is clearly wrong. The actual locality is probably in the Zuurberg mountains north of Uitenhage. This is however, only the eastern range of the species and it is not known further east than the Zuurberg Pass itself. Up here on the mountains in sour grassveld, the plants are very much like described by Baker. The leaves are glaucous, glabrous and incurved, while the stems are not unduly elongate. The full distribution of the typical species is not known. From Paardepoort in the Klein Winterhoek mountains and westwards, the species is much more variable. The stems of the plants are usually well‑developed, the leaves may be erect or spreading and are tuberculate even if only slightly so. This is the basis for recognising the var. herrei. H. glauca var. herrei fa armstrongii comprises a localised population northeast of Uitenhage ‑ based on a vegetatively propagating clone. J.R. Brown said that this form was close to ‘ H. herrei var. depauperata’ in general appearance and colour, but easily separated on the basis of having non‑confluent tubercles only on the margins and keel. The one distinguishing feature of fa armstrongii is colour ‑ both in habitat and in cultivation it tends to be darkish green and lacks the glaucous sheen of the typical species. The fa armstrongii can be superficially compared with H. fasciata except that it is caulescent. It also occurs in close proximity to H. fasciata fa browniana. Schneider (1972) showed that fa armstrongii and H. glauca were both distinguished by the absence of waxing in the stomatal chambers, and this too suggests that they belong together. In a very recent publication, Brandham and Cutler (1981), show that the fa armstrongii is a pentaploid of probable hybrid origin, showing closer affinities to H. coarctata than to H. glauca.
1999 – There is a small discrepancy in the citation of the type as Col Scott cites ‘Zuurberg Pass, Cooper sn.’ whereas Breuer and Metzing state ‘Cape without locality’. Although it may be supposed that the var. herrei is a discrete element, it is not possible to separate the herbarium specimens in this way. The var. glauca comprises the smooth eastern form, but this also occurs even west of the range for the more commonly tubercled var. herrei. The range of variation in that variety is quite considerable, as is apparent from the many species and varieties which emanated from particularly the Steytlerville area.
a. var. glauca
Distribution: 3224 (Jansenville): Fairview (-CD), Bayer 4179 (NBG); 1km N. Jansenville (-DC), Branch 321 (NBG). 3225(Somerset East): De Hoogte (-CD), Smith 3521, 5547 (NBG). 3323(Willowmore): Wilds View (-BA), Vlok 1554 (NBG). 3324(Steytlerville): Burbank (-AA), Marx 58 (NBG); 2km E. Humefield (-AA), Bayer 4168b (NBG). 3325(Port Elizabeth): Paardepoort (-AC), Archibald in NBG1320/37 (NBG), Bayer & Venter 6615 (NBG); De Plaat (-AD), Venter 91/16 (NBG); Zuurberg Pass (‑BC), Smith 12, 1183 (NBG), Marloth 4212 (PRE); Zuurberg Pass, N. Inn (-BC), Archibald 5280 (PRE).
Inadequately located: Somerset East, Smith 1184 (NBG); ex hort Ross-Frames in NBG75/44 (NBG), Britten 5748 (NBG), Henderson 1442 (NBG).
Haworthia glauca var. glauca JDV91/16 northeast of Kirkwood. It occurs here with tuberculate specimens.
Haworthia glauca var. glauca JDV90/12 Zuurgerg Pass. Without tubercles and with incurving leaves.
b. var. herrei (V.Poelln.) Bayer :122(1976). Bayer :118(1982). H. herrei V.Poelln., Feddes Repert.Spec.Nov. 26:24(1929). ibid., Cact.Succ.J(U.S.) 1930. Type: Cape, STE 5682. Not preserved. Neotype (B&M): Campherspoort, Barker 5069 (NBG): H. herrei var. depauperata V.Poelln., Feddes Repert.Spec.Nov. 31:86(1932). Type: Sundays River, between Steytlerville and Port Elizabeth, STE 6642. Not preserved: H. jacobseniana V.Poelln., Desert Pl.Life 9:102(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1038). Type: Cape, Jansenville, Mrs E. Jones in Triebn.944. Not preserved: H. eilyae V.Poelln., Kakteenkunde lO:152(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1938). Type: Cape, Steytlerville Distr., Archibald 1125. Not preserved: H. jonesiae V.Poelln., Kakteenkunde lO:152(1937). ibid., Feddes Repert.Spec.Nov. 43:109(1938). Type: Cape, Steytlerville, Mrs E. Jones. Not preserved: H. herrei var. poellnitzii Res., Mems.Soc.Broteriana: Succ.Afr. 3:89(1943). Type: Cape. Not preserved: H. eilyae var. poellnitziana ibidem. Type: Cape. Not preserved: H. eilyae var. zantneriana idem. :90 loc.cit. Type: Cape, STE6625. Not preserved. H. armstrongii V.Poelln., Kakteenkunde 10:152(1937). ibid., Feddes Rep.Spec.Nov. 43:108(1938). Jacobsen 2:537(1954). H. glauca var. herrei fa armstrongii (V.Poelln.) Bayer :99(1976). Bayer :99(1982). Type: Cape, Adelaide, Armstrong. Not preserved. Lectotype (B&M): icon (B).
herrei: for Dr.H. Herre.
None of the species or varieties described here are represented by type specimens nor good locality data, and it is perhaps fortunate that it is not very important in this case. I wrote an article for the New South Wales Study Group in 1970 which included photographs of plants collected from the Campherspoort vicinity. The variation in the clones covered easily the characteristics attributed to the elements described: narrow stems with erect incurving leaves, wider stems with erect spreading leaves, short nearly deltoid leaves, long lanceolate leaves, tubercled or non-tubercled. There has been some cytological evidence to suggest what relationship the fa armstrongii has to the species. However, the results are conflicting and it is not easy to question this (or scanning electroscopy for that matter). Brandham and Cutler (1981) report H. glauca to be ‘mostly hexaploid….but two .. were aneuploids’. Their discussion is ambivalent as on p515 of the article they suggest there is insufficient evidence to decide its parentage, whereas on p540 they state that it cannot be a variety of H. glauca because it is pentaploid and the species is hexaploid. The authors refer to two populations of H. glauca studied, but the list of materials cites only one (actually the var. herrei), and one for the fa armstrongii. Riley (1959) reported H. glauca to be tetraploid and thus a pentaploid within the species is to be expected. Finally it is now known that H. glauca is common in the area close to where fa armstrongii was originally collected. A recent visit to the site suggests that it no longer occurs there. Brandham and Cutler also looked at epidermal character where they conclude that fa armstrongii was closer to H. coarctata than to the Steytlerville collection of H. glauca. This result has to be viewed in proper perspective. It is an hypothesis based on a limited range of material and it conflicts with Schneider’s results (quoted above). Even the casual observer could probably conclude that the most logical complement to H. coarctata, both geographically and morphologically, is H. glauca. My own experience of ESM studies (and similar sophisticated techniques) is that not enough is done to first examine general variation, and that conclusions only apply within the narrow limits of the material studied.
Distribution: 3323 (Willowmore): Redcliffe (-BA), Bruyns 7053 (BOL); Miller (-BB), Acocks 19902 (PRE); 4km SW. Miller (-BB), Smith 3658 (NBG); 11km SW. Miller (-BB), Smith 3660; W. Knoetze (-BB), Smith 6917 (NBG); 11km Knoetze to Miller (-BB), Smith 6863 (NBG). 3324(Steytlerville): Campherpoort (-AA), Smith 7046, 7047, 7048, 7049, 7050, 7055, 7056, 7058, 7059 (NBG); 1km W. Campherpoort (-AA), Smith 3648, 3648a (NBG); 6km W. Campherpoort (-AA), Smith 3652 (NBG); 16km W. Campherpoort (-AA), Smith 3237, 3655, 3655a Pienaarspoort (-AB), Smith 6217, 6866 (NBG); E. Campherpoort (-AB), Smith 3592 (NBG); Waaipoort (-AD), Bayer & Bruyns 6584 (NBG); Zeekoeisnek (-BC), Bayer & Bruyns 6586 (NBG); Kleinspoort (-BD), Otzen in NBG618/41 (NBG), Smith 5170 (NBG); 3km S. Kleinpoort (BD), Smith 2924, 2924a, 2924b (NBG). 3325(Port Elizabeth): Bauerskraal (-CB), Bayer & Venter 6598 (NBG); 10km NW. Uitenhage (-CB), Smith 706, 3076, 7195 (NBG), Smith 912 (NBG,PRE).
Inadequately located: Kirkwood to Jansenville, Cunningham & Cook in NBG761/31 (BOL); Steytlerville, Taute in NBG950/37, Smith 2754; Pillans in NBG356/16, Peers in NBG68046, Malherbe in NBG489/42.
Haworthia glauca var. herrei JDV91/16 north of Kirkwood. It is the tuberculate forms which are predominant and several varieties were described from this polymorphous species.
Haworthia glauca var. herrei JDV91/121 Kleinpoort. Although supposed to have spreading leaves it does not appear as though even the geographic criterion for two varieties can be justified.
Rosette stemless, slowly proliferous, 5-7cm φ. Leaves 14-20, dark brownish-green, opaque, somewhat recurved, scabrid with small raised tubercles, margins and keel with small spines. Inflorescence slender, to 35cm. Flowers slender, tepals fused and with revolute tips.
1982 – This unusual species is the most northern record in the genus, at Groblersdal in the Transvaal. It is closely related to H. limifolia and is more prominently tuberculate. Although described as having erect leaves, in cultivation they can recurve as in H. limifolia. The spiral arrangement referred to in the original is not a distinctive feature. In H. limifolia and H. venosa the angle of leaf divergence is above 160 degrees and a twin leaf spiral is often prominent. The fact that leaves are vertically coincident in threes is purely fortuitous in such an arrangement where seven vertical parastichies will often also be detectable. It is reported to have been collected out at its original habitat, but unconfirmed reports indicate that it is known elsewhere. It is a slow-growing species which can be propagated from whole leaves.
1999 – Little further evidence is available concerning this species. Although it has obviously been collected many times by collectors since first reported, nothing has found its way into the formal record. This is a tragedy, and a repeat of the often mindless collecting frenzy in the genus Aloe during the period 1950 to about 1970. The end-product of the average collection is a small contribution to a compost heap, and good permanent record should be regarded as some tithe towards the cost of a depleted natural stock. Col. Scott also described H. mcmurtryi, which I regard as a smaller variant. The similarity to the retuse Haworthias is really fortuitous and seeking such a relationship is neither realistic nor necessary. Dr Hayashi’s suggestion (in ms.) that H. koelmaniorum is linked to H. venosa var. tessellata via a northwestern corridor is slightly more probable, but flawed for other reasons. It has a more minutely papillate and scabrous texture than H. limifolia, and also more so than H. venosa.
a. var. koelmaniorum.
Distribution: 2529 (Groblersdal): Groblersdal (‑AB), Hardy and Mauve 2267 (PRE), Hardy in KG275/70 (NBG).
Haworthia koelmaniorum var. koelmaniorum JDV88/4 south of Groblersdal. Easily grown from leaf and seed so there should be no need for reliance on field collected plants.
Haworthia koelmaniorum var. koelmaniorum JDV92/3 Groblersdal. Selection is also necessary to get speciemens which are really attractive.
b. var. mcmurtryi (Scott) Bayer comb.nov. Haworthia mcmurtryi Scott, Cact.Succ.J(U.S.) 56:69(1984). Scott :140(1985). Type: TRANSVAAL-2529(Groblersdal): Loskop, SW. Dam (-AB), McMurtry 5247 (PRE).
mcmurtryi: for Mr D. McMurtry.
This is smaller and with more prominent surface markings than the typical species. It has recently been observed and photographed by J.M. Esterhuizen at a number of sites in the Globlersdal area, and he confirms continuity with H. koelmaniorum.
Distribution: 2529 (Groblersdal): Loskop, SW. Dam (-AB), McMurtry 5247 (PRE).
Haworthia koelmaniorum var. mcmurtryi JDV93/50 southwest of Groblersdal. A smaller variety with shorter, squatter leaves.
Haworthia koelmaniorum var. mcmurtryi JDV91/7 Loskop Dam. Unlike H. limifolia the leaves can be obsurely windowed.
Haworthia koelmaniorum var. mcmurtryi JDV91/7 Loskop Dam. Also quite variable regarding tubercles and venation.