The ‘retuse’ haworthias are among the most fascinating of the genus and therefore Colonel Scott’s recent revision is of particular interest. This work…‘A revision of the genus Haworthia, Section Retusae” was published in ALOE, Dec.1973 and forms a new taxonomic record of the species involved. The object of the present paper is to examine the validity of this record particularly in the light of past history of Haworthia. A considerable literature on Haworthia is now available much of which points to the problems likely to be encountered in the group. Much of it also points to the consequences of inadequate species concepts for the group.
37. Haworthia turgida Haw., Suppl.Pl.Succ. :52(1819). Salm-Dyck, Monogr. 9:t5(1837). Bayer :163(1976). Bayer :57(1982). H. retusa sensu Scott :113(1985). Type: Cape, ex hort Kew. Not preserved. Neotype (B&M): icon, Bowie (K): H. laetivirens Haw., Suppl.Pl.Succ. :53(1819). Salm-Dyck, Monogr. 10:t3(1837). Type: Cape. Not preserved. Neotype (designated here): icon t.3, Salm-Dyck: H. caespitosa V.Poelln., Cactus J 5:33(1936). V.Poelln., Feddes Repert.Spec.Nov. 43:103(1938). Type: Cape, near McGregor, G.J. Payne in Triebn. 586. Not preserved. Lectotype (B&M): icon (B): H. caespitosa fa subplana V.Poelln. ibid. 44:232(1938). Type: Cape, Uniondale, G. Helm in Triebn. 874. Not preserved: H. caespitosa fa subproliferans V.Poelln. ibid. Type: Cape, Calvinia. Not preserved.
turgida: swollen.
Rosette partially stemless, proliferous, 5-10cm φ. Leaves 20-40, turgid often as thick as broad, recurved or slightly retused, generally mottled, yellow-green to pink in sun, margins and keel lightly spined. Inflorescence simple, 15-20cm. Flowers slender, brownish-white with darker venation.
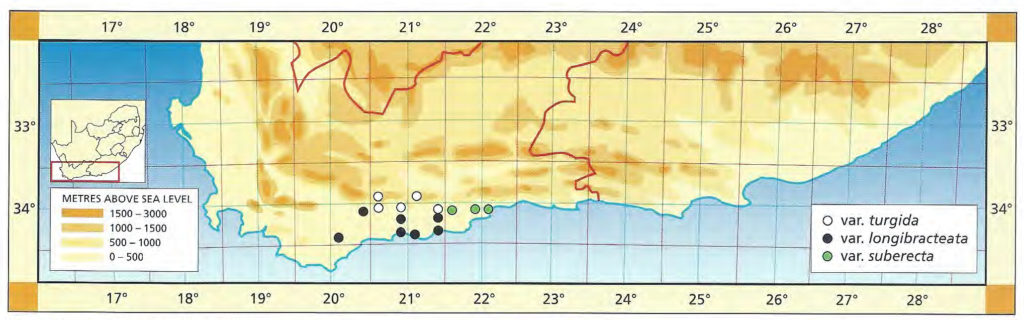
1982 – H. turgida is the most widely distributed of the Southwestern Cape species and also the most variable. It occurs in both the higher mountains in sandstone, and also on the lower lying shales. It is very subject to ecotypic variation and the best evidence of this is north of Bredasdorp eastward apparently through the coastal limestones, northward to Swellendam where there is possibly intergradation with H. reticulata, then eastwards again in the Langeberg Mountains and down all the main river valleys including the lower Breede River valley. The eastern limit is near Little Brak. The northern limit is of course the Langeberg mountain range and it is unlikely that H. turgida occurs inland along the Gouritz river ‑ despite an unconfirmed report of a collection northwest of Calitzdorp. Some of Von Poellnitz’s forms and varieties were recorded from as far afield as Calvinia and Steytlerville. Although the possibility of such collections cannot be completely ruled out, it is unlikely that these could have been relatives of H. turgida. There are forms in the Potberg mountains as well as in the Riviersonderend mountains which may intergrade with other local species (e.g. at Greyton with H. mirabilis) in the same way that H. turgida is related to H. retusa in the Heidelberg and Riversdale areas, or at Albertinia. It is a fascinating problem as it is obvious that affinities at one locality may not be the same at another. Thus it is quite conceivable that H. turgida may have an affinity with H. magnifica var. notabilis at Robertson, with H. maculata in the Hex River valley and with H. archeri through the inland mountains. H. turgida is generally a small species up to about 30mm diameter in the sandstones, however, in shales it may grow up to 80mm diameter. It is always very proliferous and grows on steep rocky slopes as opposed to H. retusa which is less proliferous and on level areas. H. turgida is also not withdrawn into the ground. This contrast of proliferation and withdrawal into the soil as opposed to clump formation also occurs in the related species H. reticulata and H. herbacea, and is evident in less strongly related species such as H. cooperi and H. cymbiformis.
1999 – The early illustrations by Salm-Dyck hardly allow room for doubt about this species, and yet the localities that von Poellnitz cites are grossly off the mark for the species and varieties which he described and cited. The problem that Col Scott had with this species and with H. retusa should also be explained here. It arises out of a misconception about leaf tiers and the statement ‘quinquefarious’ in the early literature. This term refers to vertical leaf tiers and it is apparent from a manuscript in the Grahamstown (Albany) Museum (and also from the revision in Aloe 11, 1973) that Scott concluded this to mean horizontal leaf layers. Also Scott’s treatment, where he applies the name H. retusa to this element, as well as to a number of von Poellnitz’ species and varieties H. laetivirens, is baffling. Breuer and Metzing have nominated a neotype which is not a good representation of the typical variety from the sandstones of the Langeberg mountains.
Of the southern Cape species, H. turgida is unquestionably the main role player. It occurs in recognisable form from Bredasdorp to east of Mossel Bay. It forms a continuum with H. reticulata and H. herbacea which are in the Worcester/Robertson area, and ranges from the Langeberg high peaks to the valleys running seaward and southward. It is unfortunate that apart from Smith’s names, few others have any geographical credibility. The affinities suggested in the 1982 discussion are in retrospect improbable. If there is a further association which needs exploration, it may be the link with the inland species through the high mountain forms. It does not seem sound to speculate on the chronological origins of the elements at altitude as opposed to those at lower levels.
a.var. turgida The typical variety based on Salm-Dyck’s renditions, rather than on the Kew illustration cited by Scott, which I have not seen, is considered to be the one in the Tradouw Pass east of Swellendam. The leaves are almost recurved, highly mottled and moderately spined. The higher mountain forms in the sandstones are generally smaller and note has to be taken of similarities between these smaller softer elements of apparently disparate species e.g. H. mirabilis var. consanguinea, H. maculata, and H. vlokii, all at high altitude in sandstones. At Heidelberg H. turgida expresses its full potential in terms of ecotypic variation where it passes from sandstones, to shales and on to the clays of the Witteberg series. It also appears to hybridise with both H. heidelbergensis and with H. floribunda in that area.
Distribution: 3320 (Montagu): Tradouw Pass (-BC), Read (BOL), Smith 3247, 5160, 6784, 7517 (NBG); S. Barrydale (-DC), Smith 3902 (NBG). 3321(Ladismtih): 16km N. Riversdale (-CC), Smith 7197 (NBG); NE. Riversdale (-CC), Smith 5385 (NBG). 3420(Bredasdorp): Buffeljachts (-BA), Smith 4941 (NBG); Heidelberg (-BB), Smith 5044 (NBG), Kramer in NBG841/60; N. Heidelberg (-BB), Smith 6203 (NBG); Bayer in KG240/72, in KG 241/72 (NBG); Blackdown (-BB), Smith 5546 (NBG). 3421(Riversdale): Glen (-AB), Muir 3006 (BOL); Klein Kruisriver (-AB), Smith 6075, 6080 (NBG).
Inadequately located: ex hort. Whitehill, NBG68371, NBG68056, ex hort. Ross-Frames, NBG81/44, NBG2937/32, NBG68057, NBG1827/28, Smith 3240, 3429), 3498, 3904, 3950 (NBG); Swellendam, Ross-Frames (BOL); Riversdale, Muir in NBG1827/28 (BOL); Albertinia, Muir 1300 (BOL).
Haworthia turgida var. turgida JDV89/20 north of Heidelberg. This population seems to have been influenced by H. heidelbergensis.
Haworthia turgida var. turgida JDV89/23 north of Heidelberg. A little further north the plants are transforming to the smaller typical appearance.
Haworthia turgida var. turgida JDV85/75 Tradouw Pass. It is characteristically found in the sandstones and can occur in dense masses on steep rock faces.
b.var. longebracteata (Smith) Bayer comb.nov. H. longebracteata Smith, JS.Afr.Bot. 11:75(1945). Scott :127(1985). Type: CAPE-3421(Riversdale): near Stilbaai, Dekenah 18 in Smith 5378 (NBG).
longebracteata: long bracts.
This variety is fairly large and represents the lower-lying larger forms which occur from Bredasdorp across to the Kafferkuils River east of Riversdale. The typical form is from the Stilbaai area where the leaves tend to be suberect with a fairly pronounced end-area. To the west this end-area is less pronounced and both the Bredasdorp and the Swellendam (southwest) forms are reminiscent of the flatter non-retused leaves of H. reticulata.
Distribution: 3420 (Bredasdorp): 5km SW. Swellendam (-AB), Bayer 2420 (NBG), van der Merwe in NBG1184/28, Malherbe in NBG204/41; Napky (-AB) Bruyns in KG37/77 (NBG); 20km N. Bredasdorp (-AC), Venter in NBG868/38, Malherbe in NBG470/42, Smith 3949, 5484 (NBG), Bayer in KG34/70 (NBG); Kransriviermond (-BB), Smith 5752 (NBG); Diepkloof, S. Malgas (-BC), Bayer 2533 (PRE); 3km W. Vermaaklikheid (-BD), Kramer 434 (PRE). 3421(Riversdale): Kafferkuils Bridge (-AB), Smith 6795 (NBG), Bayer 4479 (PRE); S. Riversdale (-AB), Smith 7181, 7204 (NBG); Duiwenhoksriver (-AC), Bayer 2672 (NBG); Brakfontein (-AC), Smith 6107 (NBG), Bayer in KG94/71 (NBG); Botterkloof (-AD), Smith 5378 (BOL,PRE); Kransfontein Farm (-AD), Bohnen 7735 (NBG,PRE); Stilbaai (-AD), Fouche 46 (PRE); Dekenah 18 in Smith 5378 (NBG), Smith 2242, 2811, 5382, 6794 (NBG).
Inadequately located: Heidelberg, Ferguson (BOL).
Haworthia turgida var. longibracteata JDV89/24 north of Stilbaai. Always on rocky steep slopes and in dense clusters.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Very like H. reticulata except for the larger, more turgid and spreading leaves. The flower is much less conspicuous.
Haworthia turgida var. longibracteata JDV92/30 south of Heidelberg. Becoming very colourful under stress.
Haworthia turgida var. longibracteata JDV86/6 north of Bredasdorp. A very isolated population.
c.suberecta V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, George district, Mrs Helm in Triebn. 844. Not preserved: Neotype: CAPE-3422(Mossel Bay): Brandwacht (-AA), Bayer in KG631/69 (NBG): H. turgida var. subtuberculata V.Poelln., Feddes Repert.Spec.Nov. 44:134(1938). Type: Cape, Mossel Bay, Lategan. Not preserved: H. turgida var. pallidifolia Smith, JS.Afr.Bot. 12:10(1946). Type: CAPE-3421(Riversdale): Draaihoek, Albertinia, J. Dekenah 146 in Smith 5714 (NBG).
suberecta: leaves almost erect.
Here the variety is extended to include all these more strongly retused, clump-forming populations which extend all the way from Brandwacht, northeast of Mossel Bay, to Albertinia. This variety is strongly mottled and the leaf ends are slightly truncated and rounded. Smith based his var pallidifolia on a pale-coloured variant which is actually not very common. The Gouritz River form is the most truncated and as, in the typical variety, is highly mottled with semi-translucent dots.
Distribution: 3421 (Riversdale): Draaihoek, Albertinia (-BA), J. Dekenah 146 in Smith 5714 (NBG), Smith 5045, 5490 (NBG), Bayer 4476 (PRE); Weltevrede (-BA), Smith 5537, 7202, 7203 (NBG), Bayer 4471 (NBG); Wydersriver (-BA), Smith 2890 (NBG); Droogerug (-BA), Muir 3006 (BOL); Nuweberg (-BA), Bohnen 9024 (NBG); E. Valsch River Bridge (-BA), Smith 5714, 6912 (NBG), Bayer 4477 (NBG); Gouritz River (-BB), Fouche 48 (PRE), Smith 3857, 3961 (NBG), Bayer 4478 (NBG). 3422 (Mossel Bay): Brandwacht (-AA), Herre in STE6374 (BOL), Bayer in KG631/69 (NBG)
Inadequately located: Mossel Bay, Smith 4952, 5078 (NBG); ex hort Smith 647 (NBG), Naude in NBG1
Haworthia turgida var. suberecta JDV84/74 northwest of Mossel Bay. Leaves abbreviated and rounded at the tips.
Haworthia turgida var. suberecta JDV87/108 Grouritz River. If not so proliferous this would probably be a much more popular plant.
Haworthia turgida var. suberecta JDV94/101 Brandwacht north of Mossel Bay. On stony clay slopes.
Haworthia turgida var. suberecta JDV94/88 northwest of Albertinia. The origin of Smith’s var. pallidifolia in which the leaves are very pale.
I wrote a short note for Haworthiad, to explain a picture of Breuer’s new species H. fusca (MBB7507), and said “… the fact is that it is from a small population just west of Albertinia en route to another of Hayashi’s (?) species H. esterhuyzeniae, and also to Breuer’s H. vincentii. As readers we are being conditioned to accept that there are many kinds of species such as biological species, morphological species, taxonomic species, good species, bad species etc etc. so a latin binomial could mean anything (and the word ‘tautology’ has been added to my vocabulary). Botany needs a sensible and practical handle to a squadron of populations from between Albertinia and Great Brak. I would gladly supply this if somehow I could be assured that the act was not seen to be the clown’s contribution to the circus.”
Without any assurance, but with the encouragement of Stirling Baker, I am going to try and produce an explanation.
Put very bluntly and without any apology to a group of people who definitely deserve better, my life experience is that taxonomy is largely a farce despite the fact that it works surprisingly and exceedingly well. I have already written around the subject a number of times and do not want to repeat what is not necessarily true other than the contribution these thoughts have made to my personal psyche.
In this contribution I am discuss, illustrate and then propose that there are just two species, H. retusa and H. pygmaea in a complex where presently more than nine species and varietal names are being used. I do this in consideration of all the populations of Haworthia known to me in the winter rainfall biome. Thus I recognize the need to rationalize species like H. mirabilis (which will then absorb H. maraisii, H. magnifica and H., heidelbergensis, and H. retusa (which will absorb H. turgida. There is a major problem in that the populations indicate three species in the west, viz. H. mirabilis, H. retusa and H. mutica but these appear to fuse or morph to two in the east. My past treatment of species and varieties like maraisii, magnifica, acuminata, dekenahii, argenteo-maculosa will bear witness to the nature of the (my) problem.
This chapter is based on recent field exploration and embroiders around many aspects of Haworthia species discussed in earlier chapters. What should be striking is that new populations follow the very predictable geographic pattern that all my earlier exploration has exposed and in my estimation confirm in every way what I consider a sound and satisfactory taxonomic solution and help explain its limitations.
My experience with Haworthia dates back to my childhood and on to nearly 70 years of observation. However, my interest was only able to properly manifest when I began work at the Karoo Botanic Garden in 1969 and it has since been through many phases. I wrote a formal taxonomic revision of the genus in 1999 and have spent a good bit of the last nine years adding to and verifying what I wrote. Haworthia has always been regarded as a problem child of botany to be avoided by professional taxonomists for various reasons including an apparent phobia of the many amateur collectors peering over the shoulder while at work. This has puzzled me because it seemed to me that if the need for good classification and identification was so strong there was an obligation on botany to provide the service. So my involvement has been largely by default. I was trained in an agricultural and entomological tradition with a totally different and unsophisticated approach to things like taxonomy, systematics and nomenclature. In the infant science that agriculture then was in South Africa, I can barely claim that my MSc is much more than an indication that I tried to learn something beyond normal schooling. While trained as an agricultural entomologist, my leaning was to plants and I eventually came to the Karoo garden to do what I liked best viz. exploring plants. Unfortunately the route is via identification and names and so I have walked a long road through the minefield that this is. Was this only in respect of Haworthia? No! This is a persistent misconception. Haworthia is only different because it has attracted such close and sustained amateur interest by so many for so long. I experienced failing classification in many other genera. To be fair I think the real reason is the lack of importance attached to the whole function of plant classification. It even seems as if many modern botanists pursue the study of plant relationship under the guise of systematics that is not committed to providing formal names and identifications.
In Haworthia Update 4 I wrote an essay about the haworthias east of Albertinia in which I discussed their relation to H. retusa and H. mirabilis, while generally lumping them largely in H. pygmaea. There are of course real ‘turgida’ populations as far east as near Mossel Bay, so I argued the case for an interplay of the two former species that over the whole distribution range generated two ‘species’ in the east viz. H. pygmaea and H. retusa (to include ‘turgida’), and three ‘species’ in the west adding H. mutica to H. mirabilis and H. retusa.
Recently I had the pleasure of meeting Gregory Nicholson who is studying botany at University, Cape Town. He surprised me by telling me that there was a Haworthia on his parent’s property west of Herbertsdale. It was not in fact so much surprising as confirmation of the belief I formed on a trip a short while before that there must be haworthias in the very suitable terrain of the Jakkals River valley 6km west of Herberstdale. The surprise came when Greg indicated the position of the plants much deeper into the mountains.
The writing of my grand finale was inspired by several things. One of these was another item of a mind-numbing foray into the classification of Haworthia. So I asked that deep thinker and observer, Gerhard Marx, for a devil’s advocate (abbrev. DA) point of view which he has done with the same competence he has as an artist. I have many times in my writing addressed the issue of a species definition and produced one too. Not surprisingly the first thing the DA does is dismiss my definition without producing one of his own. Simply being able to say that an indeterminate number of plants from some population are sufficiently different in respect of a character or two from other haworthias, is motivation enough for the generation of a new name?
The case of H. groenewaldii Breuer, described in an article authored in Alsterworthia 2.2:15-20 by Breuer, Marx and Groenewald is the case in point. It presents the description of this supposed new species from Buffeljags east of Swellendam. The article is written in the first person (Breuer) who quotes extensively from Gerhard’s e-mails, and includes a piece by Jannie Groenewald under the heading “Description of the Vegetation type and distribution”. The overall impression is of an article that conforms to the style of a forgotten era and it is not possible or sensible to attempt a rational dismissal. Who is actually responsible for the article and how does one correct misleading statements without giving offence?
A review of the plants photographed recently around Albertinia … I only know of 23 populations in this area that relate in different ways to the names Haworthia turgida, H. retusa, H. mirabilis, H.emelyae, H. pygmaea, H. splendens, H. vincentii, H. acuminata, H. fusca, H. dekenahii, H. argenteo-maculosa, H. pygmaea, and H. esterhuizenii. I still need to get pictures of H. splendens, H. vincentii, and H. turgida on the Wydersriver.
In essence there are two variants – the clumping cliff dwellers (H. turgida) and the mostly solitary flats dwellers (H. retusa). The flat dwellers are far more variable than the cliff dwellers. If all the variation is considered against the differences in each of the habitats, it seems very unreasonable to suggest that these populations are anything but a single species (H. retusa) expressing itself differently according to local dictates and demands. This is same issue of variability occurs in these plants all the way westwards.
97. 2020.03.04 – Esterhuizenii – Aasvoelberg. I was concerned that I am not sure anymore of flowering time because essentially retusa and mirabilis are separated by a late summer vs a spring flowering. None of the populations I saw now in late February showed any sign of buds or flower. Only the Humor plants had just two old inflorescence of about two months age. The fact it is a SYSTEM. The retusa elements are lighter coloured signified by having an element “pallidifolia”, while the mirabilis element has the element “nigra”. Recognising species as systems, variation and differences can be seen in a more realistic way. This does help to overcome the pressures that ignorance, egocentrism, emotion, commercialism and intellectualism bring to bear on classification. H. retusa ‘esterhuizenii’?
The map is useful although it does raise the question of how confidential should we be about locality. Part of this quest was to examine how populations have fared in the years since “discovery” and also in the 50 years since I have been directly involved. My observation is that the main threat is excessive animal pressure – and/or combined with drought. The Aasvoelberg population seems to be very small and a secondary threat is alien weed infestation. But I really do not know the actual size and health of the population. I did see plants further west long ago, but it will be a real mission to explore there properly. An important point about conservation is that it is really only as systems that species can cope with natural and even man-imposed change. It is only with a proper and true definition of the term “species” that we can even discuss them sensibly. “Adaptive” must surely be part of that?
98. 2020.03.05 – This is H. retusa (turgida) at Buisplaas on the Gouritz River between the Gouritz Bridge and Herbertsdale. This form was informally known as H. rodinii and is known from the Gouritz Bridge and also south of that. As a cliff dweller it is normally clumping. The first time I saw such a plant it was a specimen reputed to have come from Die Hell much further north in the Swartberg Mts. VERY unlikely, but H. retusa should be sought in the Gouritspoort where the river passes through the Langeberg Mts. There are many populations of these cliff dwellers and countless variants. I do not and cannot, dispute the fact that names present a problem for suppliers and collectors, but that is not a problem that can be settled by science. Association of a botanical binomial and place is the only solution that makes sense to me. Taking out the geographical detail diminishes the sense of any name. But this series is to explain and expand on this issue.
These are also Buisplaas but in a totally different habitat several hundred meters from the cliff dweller. One picture shows a very proliferous clone. This same juxtaposition of clumping and non-clumping occurs elsewhere and is a pronounced feature of the H. cooperi and H. cymbiformis problem in the Eastern Cape. These Buisplaas populations validate my conviction that the Western cape retusoids are in fact a single species system. There has been a predictive element in all my exploration since the revision of 1999 (Haworthia Revisited) and this population simply fits a pattern. Comparing all the pictures of this series I am now showing will demonstrate this too. H. retusa ‘pygmaea’. I write the name like this because in the absence of a species definition, formal lower ranks have to be meaningless. Furthermore the nature of local variation is such that it is generally impossible to circumscribe ranks. A very good example of this is the persistent reference to H. mirabilis var magnifica which can absolutely not be supported on the basis of a type specimen representing an original group of such similar or like plants.
99. 2020.03.07 – Despite the drought and animal activity (and human predation presumably) – the Cooper Siding plants are in great health. I am posting all my pictures (of Haworthia – there are many other plant species of significance present) because the reality of diversity within populations gets zero attention in citizen science and little more in academia. But do not get me wrong. I write “citizen science” not citizen cognition. Also I do not ignore the fact that “citizen science” also features in academia. These plants (a plant?) was described as H.dekenahii var argenteo-maculosa. In my Handbooks 1976, 1981), I placed the two under H. retusa. Was this wrong? No – because moving them to H. pygmaea as I did in my revision (1999) highlights the reality that this is a single system and not just spatially isolated populations that can be treated as different.
In writing about Albertinia and the conservation state of these Haworthia populations (that really bothers me), some very serious thoughts come to mind. Do we really want to know the problem and the answer(s)? The problem is environmental deterioration – climate change. This is coupled with the decline in moral values across the counter. But do we want to acknowledge this and really want to know what is true. That all of existence is actually a grand illusion that we may choose to be locked in to. I should not have suggested that climate change is the main factor – although Albertinia has been experiencing the worst drought in living memory. Livestock pressure also definitely impacts negatively on vegetation, and our dietary addictions prevent us recognizing this fact.
100. 2020 03.08 – Dekenahii is at Draaihoek on the Valse River northeast of Albertinia. Juxtaposed again on a nearby rocky cliff is “turgida pallidifolia”. A floribunda / chlorocantha variant is in close proximity in its own preferred space. A different form of the turgida retusoid used to be on north facing cliffs a little further north at Weltevrede. Keep in mind that I am considering a bigger picture in which differences within populations negate differences between them, and especially when habitat is considered. I also juggled Dekenahii between retusa and pygmaea and whatever you do with it, the best explanation I can offer as a binomial is H. retusa ‘pygmaea’. Better still is to add the unwritten meaning that it is actually Helgaard Oosthuizen’s discovery of H. retusa as a group of plants at Draaihoek circa 1943 given to Smith by Dekenahii. Names standing alone may be good for business but the interest lies in the information attached to them and which they point to.
This is the “turgida” form of H. retusa at Draaihoek. These plants have more pointed leaves and are more colourful than the “pallidifolia” that I remember 59 years back. But the precise locality may have been different. I remember plants more like those at Tweekuile further east along the river that I will still post pictures of. At Weltevrede further upstream the plants had longer leaves. Steven Molteno coincidentally reminded me of H. marumiana and the exact same conundrum I am trying to explain here, but magnified manifold over the vastness of the Karoo and our ignorance of both marumiana and the Karoo.
101. 2020.03.10 – Tweekuile is north of Albertinia and just a few km east of Draaihoek on the Valse River. This is what H. retusa ‘turgida’ looks like there. It occurs to me that sensible taxonomy, like prevention and cure of disease, is not a sustainable business model 😅
Without any simple and practical measure, how does one deal with an endless array of slightly different looking things affected by age, exposure to direct sun and an also seemingly endless lot of agencies affecting appearance. Compare these two Cooper Siding and Humor pygmeas with the Tweekuile turgidas.
102. 2020.03.11 – Have a good look at the rest of Humor. I suppose it is still fashionable to dismiss variation like this as “hybrid swarm”. Problem here is what parents where and when. To be strictly correct, is there really any option but to say H. retusa ‘pygmaea”?