8. Haworthia cooperi Baker, Saund.Ref.Bot. 4:t.233(1871). Bayer :109(1976). Bayer :33(1982). pp. Scott :103(1985) as to Adelaide. H. arachnoidea (Haw.) Duv. sensu Pole-Evans, Flow.Pl.S.Afr. 7:t.248(1927). Type (B&M): Cape, Cooper (K): H. vittata Baker, loc.cit. :t.263. Type: Not preserved. Lectotype: icon t.234, Saund.Ref.Bot. cooperi: for Thomas Cooper.
Rosette to 120mm φ, often proliferous, stemless. Leaves 20-40, fleshy, swollen, oblong-lanceolate, quickly tapering, acuminate or truncating, marginal spines <2mm long if present. Bluish-green in colour, slightly translucent, with veins usually reddening and leaves developing purplish hues in exposed situations. Inflorescence compact, firm peduncle with many closely arranged flowers, to 20cm long. Flowers 20-30, perianth white.
1982 – This species is beautifully illustrated in Refugium Botanicum and two other species, H. pilifera and H. vittata, were also described and illustrated here. The name H. pilifera has been most commonly used for plants with relatively blunt leaves which occur in the Eastern Cape particularly around King Williamstown. The name H. vittata was similarly applied to the longer-leaved forms in the Thomas River and Cathcart areas which probably intergrade with H. bolusii. However, the name H. cooperi can equally be applied and has page preference over the other two names. H. cooperi is very common widespread and variable as the synonymy suggests. It is logical to suppose that in this case Haworth must have received specimens at some time or another. Uitewaal attempted to apply the name H. obtusa Haw. which has since been refuted by Bayer and Pilbeam (1974). Scott similarly implemented H. altilinea Haw. but uses the name in a restricted sense to exclude all three of Baker’s names, as well as names such as H. limpida and H. mucronata. The writer’s contention is that the name H. altilinea is a source of confusion and should for the present at least, be rejected. There are thus the four species with translucent bluish-, or greyish-green leaves as indicated in the key. H. cooperi is distinguished by its generally less hairy or shortly haired leaves. The leaves are obtuse to obtuse-ovate although forms with longer, more acuminate leaves also occur. These usually have thicker leaves than corresponding forms in H. bolusii and the leaves are also less hairy. Scott comments on the withdrawal of plants into the ground, and the question of exposure and soil substrate are responsible for much of the variability in the species. In ‘stayneri’ collected at Bethelsdorp near Port Elizabeth, and in several other populations, the leaf veins actually become necrotic and the leaf-ends truncated. In ‘gordoniana’ from Hankey this does not occur.
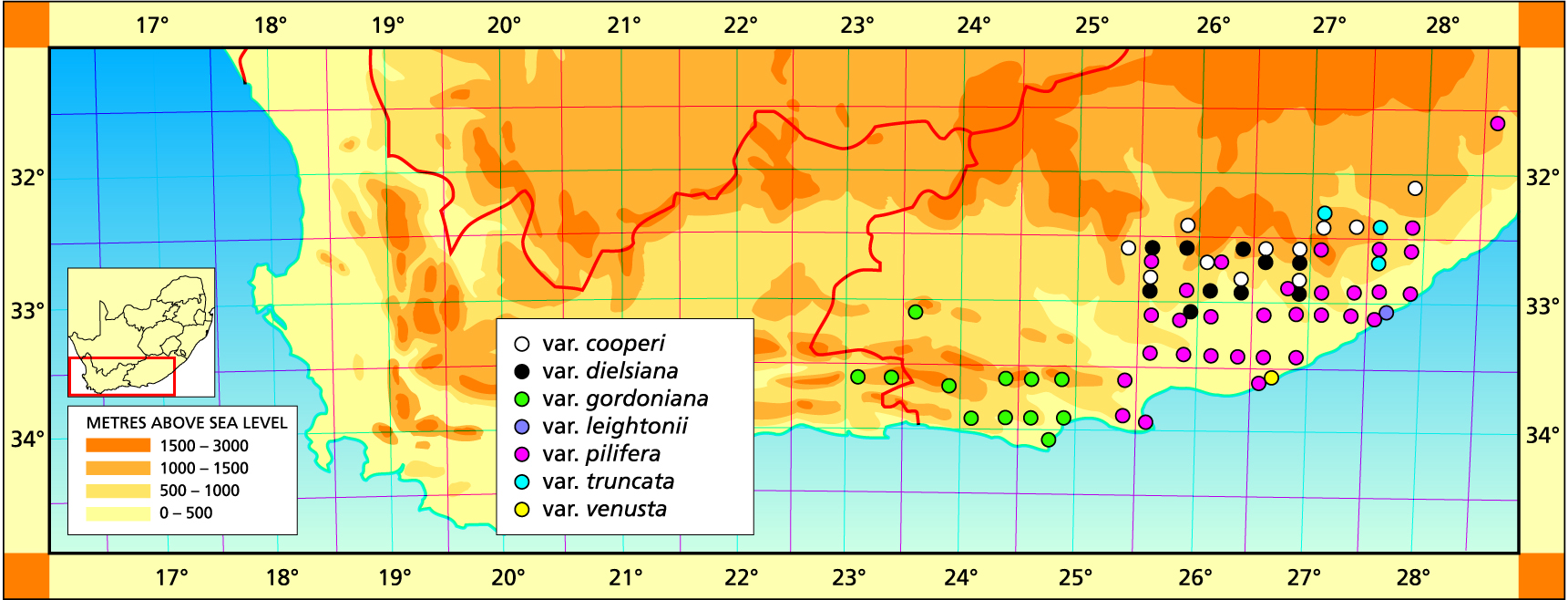
H. cooperi is a species of the dry grassveld areas of the Eastern Cape and occurs in the high-lying sourveld of the Zuurberg to the dry Valley Bushveld of the Fish River. Size varies enormously from small glabrous plants less than 40 mm in diameter southwest of Grahamstown, to enormous hairy specimens over 100 mm in diameter at Keiskamma. The variety leightonii is exceptional in that it occurs on the edges of exposed granite slabs west of East London. This variety is very proliferous and is characterised by the persistence of reddish coloration in the leaf veins.
1999 – One of the difficulties with this species is in its relationship with H. bolusii var. blackbeardiana, and here the typical variety is applied to that intermediate element in the Thomas River area. This does not represent the main body of the species at all, which is a product of the nomenclatural system. Also the previous Handbooks do not present any idea of the variation in this species. There is an additional problem in the association with H. cymbiformis. Uitewaal really exposed the problem with his recognition of the name ‘obtusa’ of Haworth for this species. Bayer and Pilbeam refuted this by suggesting that Uitewaal had misinterpreted Haworth’s description and the illustration associated with it. Scott similarly refuted Uitewaal and also applied the name as a variety of H. cymbiformis. However, he illustrated a blue-green variant which we regard as H. cooperi var. obtusata, a new name, in this work. There is clearly a problem in finding a point of origin for ‘obtusa’ as variant of H. cymbiformis rather than of H. cooperi, and that is answered under the former species. However, herbarium records indicate a problem in the broader Alicedale/Adelaide areas where truncation and abbreviation of the leaves of both species seems to be evident, together with the problem of green versus blue coloration. The essential difference between the two species – which generally have the same distribution range and very often co-occur – is that H. cooperi is an open ground species, whereas H. cymbiformis occurs on rocky shelves and cliffs. It is interesting that even a variety like leightonii has a tendency to resemble H. cymbiformis, by acquiring an opaque yellowish-green coloration as opposed to the required blue-green for the species cooperi. This is a common problem in trying to absolutely circumscribe the species. But even outside of that, there has been a massive problem in the plethora of names attached to either species. Both extend into the mountains north and west of Port Elizabeth and it is not clear just how this relates to other species which do the same. The following varieties are recognised.
a. var. cooperi.
The typical variety occurs in the Thomas River area and is along the transition zone to H. bolusii var. blackbeardiana. The distinction is that the leaves are slightly more acuminate and the venation tends to acquire reddish tints, while general coloration tends to a purplish hue. The leaf spines are also generally less than 2mm long.
Distribution:
3225 (Somerset East): 30km S. Cradock (-BD), Smith 5194 (NBG); Bruintjieshoogte (-CB), Britten in PRE 34923, Bayer 2024 (NBG); 11km S. Somerset East (-DC), Smith 2841 (NBG). 3226 (Fort Beaufort): Bobbejaanrivier, Bedford (-CA), Smith 2244 (NBG); Koonap Bridge (-CD), Bayer & Bruyns 6563 (NBG); Elandskop, Adelaide (-CD), Smith 2687, 2797, 2799 (NBG); Adelaide (-CD), Krynauw in NBG270/43; Katberg (-DA), Herre in STE6607 (BOL), Read in BOL71291, Smith 2773 (NBG); Warfield (-DB), Venter 91/82 (NBG); Woburn (-DB), Smith 578 (NBG); Alice, Stewart Memorial (-DD), Smith 5211 (NBG). 3227 (Kingwilliamstown): Thorn River near Cathcart (-AC), Acocks 11003; W. Cathcart (-AC), Bayer in KG 392/70 (NBG); Thomas River (-AD), Smith 3631 (NBG), Scott 1720 (NBG), Stayner in KG402/61 (NBG); Tsomo (-BB), Branch 13 (NBG).
Inadequately located: ex hort. Stanford (BOL).

Haworthia cooperi var. cooperi JDV91/26 Koonap River. On a south-facing rocky slope. 
Haworthia cooperi var. cooperi JDV91/98 Thomas River. In the Cathcart area there are also plants with less of the purplish colouration and which may also be without spines – confused with H. bolusii var. blackbeardiana. 
Haworthia cooperi var. cooperi JDV91/26 Koonap River. Note the darker lines and purplish tints. 
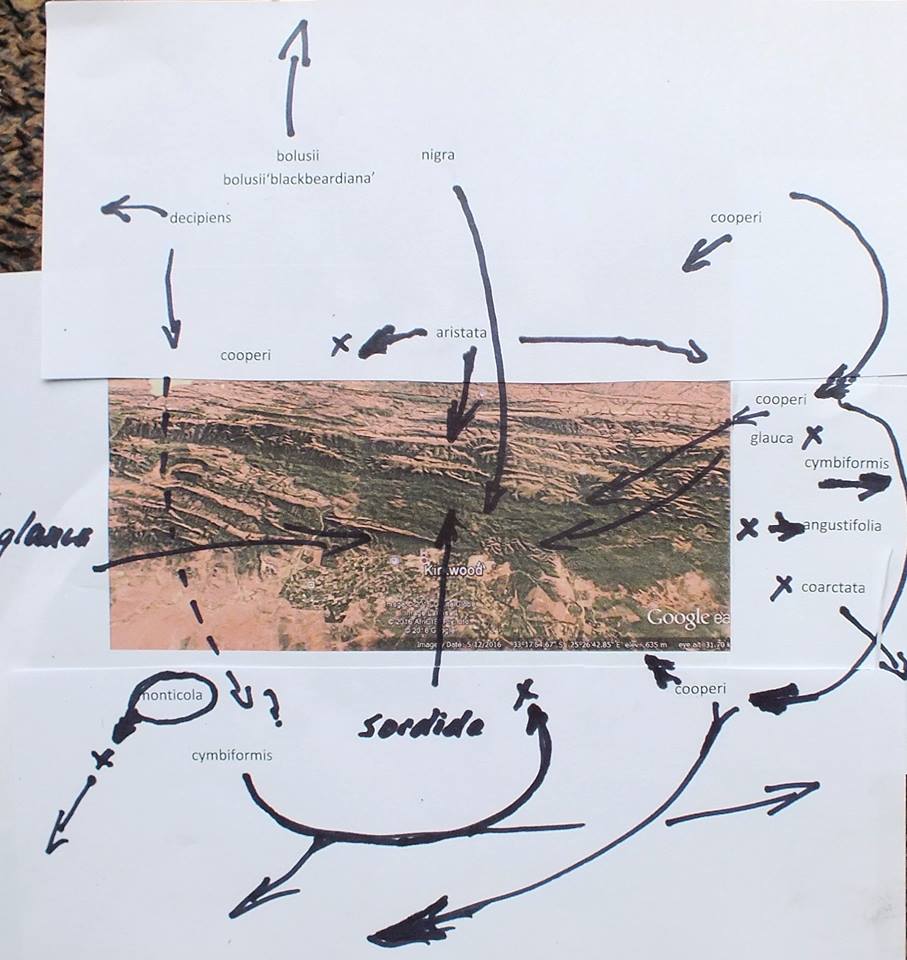
Haworthia cooperi var. cooperi JDV91/14 De Plaat, Kirkwood. Here there may be an interaction with H. gracilis.
b. var. dielsiana (V.Poelln.) Bayer comb. nov.
H. dielsiana V.Poelln., Feddes Repert.Spec.Nov. 28:103(1930). H. pilifera var. dielsiana idem. 49:27(1940). H. obtusa var. dielsiana (V.Poelln.) Uitew., Succ. 29:50(1948). Type: Cape, Sheldon, H.H. Hutton 489. Not preserved. Neotype (designated here): CAPE-3325(Somerset East): Sheldon (-BB), A.J. van der Merwe in Smith 1140 (NBG). H. joeyae Scott, Bradleya 13:80(1995). Type: CAPE-3225( Somerset East): a few kilometers NE. of Cookhouse (-BD), Scott (GRA).
dielsiana: in honour of Prof. Diels of Berlin, Dahlem.
There appear to be two elements within the species with very truncated leaf ends. The first is this variety from the western area in which the leaves are variously truncated and obtuse. Smith records a wide range of forms with or without leaf spines and some with very rounded leaf-ends. Sometimes the leaf-margins are almost ridge-like. The leaf-tips do not appear to become necrotic as happens in the var. pilifera. The leaf awn is virtually absent and the leaves tend to have marked venation. The keel and margins tend to be rounded.
Distribution:
3225 (Somerset East): Sheldon (-BB), Smith 1140 (NBG), Hutton in Herre 489 (BOL); 5km E. Somerset East (-DA), Bayer & Bruyns 6565 (NBG); Glen Avon (-DA), Smith 5790 (NBG); Merantes Kloof (-DB), Smith 3492 (NBG); Eastpoort (-DB), Smith 3491, 3493 (NBG), Reynolds (BOL), Bayer & Bruyns 6558, 6559, 6560 (NBG); Little Fish (-DC), Smith 5550 (NBG). 3226 (Fort Beaufort): 10km N. Adelaide (-CB) Krynauw in NBG267/43, Krynauw in NBG687/41; 19km NE. Adelaide (-CB), Krynauw in NBG269/43 (NBG); Chancery Hall (-CB), Bayer & Bruyns 6564 (NBG); Kagasmond (-CC), Krynauw in NBG273/43; Paardefontein, SE. Adelaide (-CD); Brakfontein, Kroomie (-CD), Smith 5112 (NBG); Koonap (-CD), Smith 7131 (NBG); Adelaide (-CD), Smith 2687 (NBG); 30km S. Adelaide (-CD), Smith 2240 (NBG); Somerset East (-DA), Fouche in PRE 34919; Woburn; Tyumie river (-DB), Acocks 13575 (PRE); W. Fort Beaufort (-DC), Bayer & Bruyns 6592 (NBG); S. Fort Beaufort (-DC), Bayer & Bruyns 6591 (NBG); E. Fort Beaufort (-DC), Smith 3489 (NBG); 16km N. Cradock road (-DC), Britten in PRE 34918.
Inadequately located: Ross’ Mission, Smith 7492 (NBG); Zaysdorp NBG1019/25; ex hort Whitehill (NBG), Ross-Frames (NBG), Stanford (BOL).

Haworthia cooperi var. dielsiana JDV91/101 Eastpoort. This photograph is of a plant from south of Eastpoort, where they are slightly greener and the leaves less truncated than in the type of ‘H. joeyae Scott’. 
Haworthia cooperi var. dielsiana JDV91/101 Eastpoort. This is a plant with very truncated leaves with virtually no end-awn.
c. var. gordoniana (V.Poelln.) Bayer comb. nov.
pp H. cooperi (Bak.) Bayer :119(1972). pp. Bayer :33(1982). H. gordoniana V.Poelln., Feddes Repert.Spec.Nov. 42:269(1937). H. pilifera var. gordoniana V.Poelln. idem. 44:237(1938). H. obtusa var. gordoniana (V.Poelln.) Uitew., Succulenta 29:50(1948). Type: Zuurbron, Hankey, Long 814. Not preserved. Neotype (designated here): CAPE-3324 (Steytlerville): Patensie (-DD), Smith 3028 (NBG).
gordoniana: in honour of Gordon King.
There are many variations of small glabrous and spined plants in the Hankey/Patensie area and it is rather problematic to decide to which species they really belong. Von Poellnitz associated his species with H. cooperi as recognised in this work and it is thus the small blue-green element which is nearly identical to his H. stayneri. Unlike the latter which is made synonymous with H. cooperi var. pilifera the leaf tips do not truncate on exposure to direct sun. It differs from similar plants which are regarded as H. gracilis, in that the outer leaves stay incurved and are sparsely spined. Also the leaves are usually more thickly turgid than in that species. The plants are less proliferous and tend to be withdrawn into the soil.
Distribution:
3323 (Willowmore): Redcliffe (-BA), Bruyns 7062 (BOL); Uniondale Poort (-CA), Bayer 4404 (NBG); Damse Drif (-CA), Bruyns 1651, 1652 (NBG); Nuwekloof (-CA), Bruyns 1840 (NBG); Saptou (-CB), Bruyns 7079 (BOL); Studtjes (-DB), Bruyns 2190 (NBG). 3324 (Steytlerville): Enkeldoorn (-CB), Perry 1427 (NBG), Bean (NBG); Holgat Kloof (-CC), van Jaarsveld 6885 (NBG); Moordenaarskloof (-CD), Stayner in KG673/71 (NBG); Grootriver Poort (-DA), Bruyns 2202 (NBG); Quagga to Cambria (-DA), Smith 2912 (NBG); Cambria (-DA), Smith 2904 (NBG); Ouplaas (-DB), Bruyns 7043 (BOL); 2km E. Hankey (-DD), Rossouw 141 (NBG); Patensie (-DD), Smith 3025 (NBG); E. Patensie (-DD), Smith 2908 (NBG); S. Hankey (-DD), Smith 3186 (NBG); Hankey (-DD), Smith 2905 (NBG); 5km N. Hankey (-DD), Bayer in KG193/73 (NBG); N. Hankey (-DD), Smith 2883, 3687 (NBG); NE. Hankey (-DD), Bayer 4474 (NBG), Stayner in KG180/71 (NBG); E. Hankey (-DD), Smith 2597, 2893, 2980 (NBG); Stayner in KG180/71 (NBG); 7km N. Zuurbron (-DD), Smith 3671 (NBG); 5km N. Zuurbron (-DD), Bayer & Bruyns 6553 (NBG); 8km N. Zuurbron (-DD), Bayer & Bruyns 6554 (NBG); N. Joubertina (-DD), Venter 91/64 (NBG). 3325DC Coega Kop(-DC), Long 1132 (PRE). 3424 (Humansdorp): Jeffrey’s Bay (-BB), Bayer & Venter 6597 (NBG).
Inadequately located: Herre STE6609 (BOL).

Haworthia cooperi var gordoniana JDV91/8 Baviaanskloof. A smaller plant with non-truncating leaf-tips and often confused with H. gracilis. 
Haworthia cooperi var gordoniana JDV94/76 north of Humansdorp. The bluish-green colour is usually a distinguishing character, and the rosettes are fairly globose. 
Haworthia cooperi var gordoniana JDV96/92 Jeffrey’s Bay. Here the plants in the field are brownish and may be confused with H. gracilis.
d. var. leightonii (Smith) Bayer
:128(1976). Bayer :34(1982). H. leightonii Smith, JS.Afr.Bot. 16:10(1950). Scott :107(1985). pp. H. cooperi Bak., Scott, Cact.Succ.J(U.S.) 53:70(1981). Type: CAPE-3327 (Peddie): Kayser’s Beach (-BA), Smith 6938 (NBG).
leightonii: for I. Leighton.
There are several populations of this variety and the circumscription is enlarged to include those from further to the northwest in which the leaves are also more lanceolate and untruncated. It is almost certainly an ecotype associated with the granitic slabs of the coastal area near Kayser’s Beach, where the plants are very proliferous indeed. This, and the strong purplish coloration, characterise the variety.
Distribution:
3327 (Peddie): Kaysers Beach (-BA), I.H. Leighton in NBG 68362, Smith 6938 (NBG), Bayer in KG6/72 (NBG), Venter 91/105 (NBG); SW. Paynes Hill (-BA), Smith 514 (NBG); Bayer 1621 (NBG).
Inadequately located: Kingwilliamstown, Taylor 3039 (NBG), Leighton in NBG662/34.

Haworthia cooperi var. leightonii JDV91/105 Kyser’s Beach. It is also very proliferous. 
Haworthia cooperi var. leightonii JDV91/97 east of Chalumna. This variety is characterised by the strong reddish-pink colour which develops under stress, but here the plants are greener. 
Haworthia cooperi var. leightonii JDV91/104 Kayser’s Beach. Associated with granite slabs. 
Haworthia cooperi var. leightonii JDV91/105 Kayser’s Beach. A population in the area has plants which bear some similarity to H. cymbiformis.
e. var. pilifera (Baker) M.B.Bayer comb.nov.
H. pilifera Baker, Saund.Ref.Bot. 4:t.234(1871). V.Poelln., Feddes Repert.Spec.Nov. 44:236(1938). Scott :104(1985). H. obtusa var. pilifera (Baker) Uitew., Succulenta 29:50(1948). Type: Not preserved. Lectotype (here designated): icon, :t.234, Saund.Ref.Bot.: H. stayneri V.Poelln. loc.cit. 42:270(1937). H. pilifera var. stayneri idem. 44:237(1938). H. obtusa var. stayneri (V.Poelln.) Uitew. loc.cit. Type: 14m Port Elizabeth to Uitenhage, F.J.Stayner. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): 13.8m Port Elizabeth to Uitenhage (-DD), F.J.Stayner in KG 2/70 (NBG): H. stayneri var. salina V.Poelln., Feddes Repert.Spec.Nov. 42:271(1937). H. pilifera var. salina idem. 44:237(1938). H. obtusa var. salina (V.Poelln.)Uitew. loc.cit. Type: Cape, Bethelsdorp Salt Pan, Mrs King. Not preserved. Neotype (designated here): CAPE-3325 (Port Elizabeth): Bethelsdorp salt pan (-DD), Smith 5817 (NBG): H. pilifera var. dielsiana fa acuminata V.Poelln. loc.cit. 49:27(1940). H. obtusa var. dielsiana fa acuminata (V.Poelln.) Uitew. loc.cit. Type: 4m from Kingwilliamstown, F.A.Fouche. Not preserved. Neotype (designated here): CAPE-3227(Kingwilliamstown): 5km Kingwilliamstown to Pirie Dam (-CC), Smith 3913 (NBG): H. altilinea Haw. sensu Scott, Natn.Cact.Succ.J 34:53(1979). Scott :84(1985).
pilifera: with hairs.
This variety constitutes the main body of the species which is centred about Kingwilliamstown and Grahamstown. It is characterised by the obtuse acuminate end-area with a pronounced point to the leaf. The margins and keel are sharply angled. It is very closely drawn into the soil surface and, on exposure to direct sunlight truncates to form a necrotic end-area.
Distribution:
3128 (Umtata): Viedgesville (-DA), Rush in KG87/80 (NBG). 3225(Somerset East): Little Fish (-DA), Smith 5551 (NBG); Cookhouse, Patryshoogte (-DD), Long 1462 (PRE). 3226 (Fort Beaufort): 24km S. Bedford (-AC), Bursey in KG335/70 (NBG); Good Hope, Alice (-DD), Smith 5496 (NBG); Fort Hare, Cressey in NBG2132/26 BOL); Alice (-DD), Smith 5210 (NBG); Middledrift (-DD), Smith 5342 (NBG). 3227 (Kingwilliamstown): Peddie (-AA), Smith 5654 (NBG); Fort Murray Bridge (-BD), Bayer (NBG); E. Kieskammahoek (-CA), Smith 6065 (NBG); Kieskammahoek (-CA), Smith 5309, 7350 (NBG); Stayner (NBG); 5km Kingwilliamstown to Pirie Dam (-CC), Smith 3110, 3913 (NBG): (-CC), Dyer 2079 (PRE); Golf course (-CD), Scott 1980 (PRE), Crampton in NBG1096/28 (NBG), Taylor in NBG250/34 (NBG); Line Drift (-CD), Smith 5434 (NBG); Balazi (-CD), Smith 3574 (NBG); St Johns Drift (-DA), Bayer 1623 (NBG); Near Komgha (-DB), Flanagan 1116 (BOL, PRE); Brigadoon (-DC), Bayer 4460 (NBG); Buffalo River (-DC), Smith in NBG343/35 (NBG); Bridal Drift (-DC), Smith 386 (NBG); Cambridge (-DD), Grenfell in NBG872/35. 3325 (Port Elizabeth): Cookhouse to Zuurberg (-BA), Smith 3494 (NBG); Shenfield (-BB), Blackburn in BOL71305; Kommadagga (-BB), Bruyns 1651 (NBG); 18km N. Zuurberg Inn (-BC), Stayner in PRE 57676; Enon (-BC), Dyer 498a (PRE); Zuurberg (-BC), Stayner in KG685/71 (NBG); Addo Park (-BD), Branch 31 (NBG); Bauerskraal (-CB), Bayer & Venter 6559 (NBG); Despatch (-CD), Muir 13133 (PRE). 1km W. Uitenhage (-CD), Smith 5818 (NBG), Britten (BOL); 22km Port Elizabeth to Uitenhage (-DC), F.J.Stayner in KG 2/70 (NBG); E. Uitenhage (-DC), Smith 3582 (NBG); Bethelsdorp salt pan (-DC), Smith 5817 (NBG), Stayner in KG2/70; Redhouse (-DC), Paterson 431 (BOL). 3326(Grahamstown): E. Riebeek East (-AA), Smith 5219 (NBG); Willowfountain (-AA), Bayer 1622 (NBG); Blaauwkrantz (-AC), Dyer 2306 (PRE); NW. Salem (-AD), Bayer 4450 (NBG); 8km N. Grahamstown (-AD), Smith 5628 (NBG), Britten in BOL71290; The Fort (-BA), Smith 5065 (NBG); Keiskamma near Breakfast Vlei (-BB), Acocks 11871 (PRE); Debe Nek (-BB), Smith 3545 (NBG); N. Committees (-BB), Smith 5435 (NBG); E. Committees (-BB), Smith 5430 (NBG); Committees (-BB), Smith 2428, 5352, 5353, 5366 (NBG), 16km N. Cradock road (-BC), Marloth 12607 (PRE); Grahamstown (-BC), Whitmore in NBG1238/24 (NBG), Erens in PRE 34929; Bothas Hill (-BC), Smith 5355 (NBG); Frazers Camp (-BD), Smith 5215, 7413 (NBG); Peninsula (-DA), Bayer in KG383/70 (NBG); Brigadoon (-DC), Bayer in KG382/70 (NBG). 3327(East London): S. Kingwilliamstown (-AB), Krynauw in NBG684/41 (NBG); E. Chalumna (-BA), Smith 5773 (NBG).
Inadequately located: Cape ex hort, Marloth 5861 (PRE); Grahamstown, Dyer 1 (BOL), Britten in BOL71289; Leighton in NBG662/34, in BOL71301 (BOL); Kingwilliamstown, Sim 1030 (BOL), Herre in STE6619 (BOL); Mquakwebe, Leighton in BOL20734-6, in Smith 6938 (BOL).

Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. Not very proliferous and drawn into the soil where they are cryptic when it is dry. 
Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. Growing very near a population of H. gracilis, and the difference is in the way this species has shorter stumpier leaves. 
Haworthia cooperi var. pilifera JDV94/96 north-east of Grahamstown. The stumpy leaf-ends may be attractively veined and may become almost necrotic to exclude light.
e. var. truncata (Jacobs.) M.B.Bayer comb. nov.
H,. obtusa var. truncata Jacobs., Nat. Catt. Succ. J. 10: 81 (1955). Type: Not preserved. Neotype: (B&M in ms.): icon in Jacobsen, Handbuch der Sukk. Pfl. 724, f644 (1956). Epitype: CAPE-3227 (Stutterheim): Runlets, Mgwali (-DA), Smith 5295 (NBG).
truncata: with obtuse leaves.
Rosette proliferous, to 70mm φ, Leaves 20-25, 20-25 x 8mm wide, pale blue-green, erect, truncated, translucent and lightly veined above. (A var. cooperi foliis parvioribus truncatis differt).
I am grateful to I. Breuer for drawing attention to the possible correct application of this name. The important arbiter would actually be colour, which Jacobsen does not stipulate. However, Breier suggests the plants from the Bolo Reserve distributed under I.S.I. No. 1762 represent Jacobsen’s variety; otherwise I would have incorporated it with H. cymbiformis var. obtusa. This is a smaller variety than the western var. dielsiana with similarly truncated leaves, and the distinction is largely on geographical grounds. Other differences are the smaller size, the rapid off-setting, and the almost unlined leaves of the var. truncata as opposed to var. dielsiana. The var. truncata is at the northeast of the distribution range of the species where it extends into the Transkei. Both Bayer and Pilbeam, and Scott have attempted to refute Uitewaal’s interpretation of H. obtusa Haw. as an earlier synonym for H. cooperi. This is a case in which the option is entirely an open one. How and why Uitewaal chose to depart from the general application of the name seems to be the norm for Haworthia.
Distribution:
3227 (Kingwilliamstown): Hunts Drift (-AC), Smith 5175 (NBG); Inerbolo (-BC), Bruyns (NBG); Mgwali (-BC), Smith 5193 (NBG); Runlets, Mgwali (-DA), Smith 5295 (NBG).

Haworthia cooperi var. truncata MBB 386/70 Mgwali, Sutterheim. Grows in a level rocky but grassy area, and forms large blue-green clumps.
g. var. venusta (Scott) Bayer comb.nov.
H. venusta Scott, Bradleya 14:87(1996). Type: CAPE-3226 (Grahamstown): NE. Alexandria (-DA), Britten 781 (GRA).
venusta: charming, and for Miss Grace Britten. Her excellent herbarium sheet led to the recent rediscovery by Gerhard Marx.
This very handsome variety is, surprisingly, coarse white-haired, exaggerating the slightly hairy tendency foundin the other varieties. It offsets sparsely if at all, and remains quite small.
Distribution:
3326 (Grahamstown): NE. Alexandria (-DA), Britten 781 (GHS, NBG), Britten and Archibald 781 (BOL, PRE).

Haworthia cooperi var. venusta JDV94/5 south of Grahamstown. A really remarkable and dramatic variety on account of the short shiny hairs. 
Haworthia cooperi var. venusta JDV94/5 south of Grahamstown. Known only at one locality. It seems to set seed easily and should become a favorite in collections.