
12. Haworthia floribunda V.Poelln., Feddes Repert.Spec.Nov. 40:149(1936). idem. 44:228(1938). Bayer :117(1976). Bayer :39(1982). Scott :58(1985). Type: Cape, Heidelberg, Mrs E. Ferguson. Not preserved. Lectotype: icon (B). Epitype (B&M): Blackdown, N Heidelberg, Bayer 158 (NBG).
floribunda: many flowered.
Rosette stemless, up to 3cm φ, slowly proliferous. Leaves 20-30 dark green opaque, upto ovate-lanceolate, spreading, twisted with flattened, rounded tip, margins scabrid to dentate. Inflorescence simple, to 250mm. Flowers 10-15, greenish-white, few open together.
1982 – This is a very interesting small species with twisted lanceolate leaves with blunt rounded tips. It was described from plants collected north of Heidelberg (Cape) where they are all glabrous and where hybridisation with H. turgida also occurs. It has not been collected further west although there is a very old collection in the Botanical Research Institute (PRE) from Swellendam. Around Riversdale the plants may have scabrous leaves with denticulate leaf margins. There is a known population north of Albertinia in which the plants have more and shorter leaves, as well as another similar population near Gouritzmond. These two populations may suggest an affinity with H. chloracantha, as put forward by A.E. Speechley (unpublished). There may be such a relationship, but it seems likely that H. floribunda and H. parksiana are in fact related. They both tend to grow well‑shaded and in moss and lichen. Further exploration of the Gouritz River tributaries may produce an answer to this puzzle.
1999 – The Gouritz valley has provided no answers to the above puzzle but there have been a number of other significant collections. There is a population south of Swellendam of which specimens in cultivation have been very robust. They are relatively light green in cultivation and also individual plants are smooth as was the original type. Odd specimens have pointed leaves. Northwest of Swellendam is a similar but darker green plant also with pointed leaves and this is taken to be H. variegata. Further south from Swellendam, H. floribunda assumes the same form as around Riversdale. At Great Brak there is a population of plants growing with H. pygmaea which were assumed to be H. floribunda, but they are probably simply H. chloracantha var. denticulifera. This is true also of the populations south of Albertinia and also of one population south of Heidelberg which has pointed leaves. The flattened leaf-tip is not confined to this species, and appears in H. magnifica and H. maraisii. A relationship with H. variegata is probable through the two populations in the vicinity of Swellendam.
a. var. floribunda.
The typical variety is not typical of the species at all and this glabrous variety is only known from the type locality at Heidelberg where it is very scarce. Hybrids with H. turgida are also present.
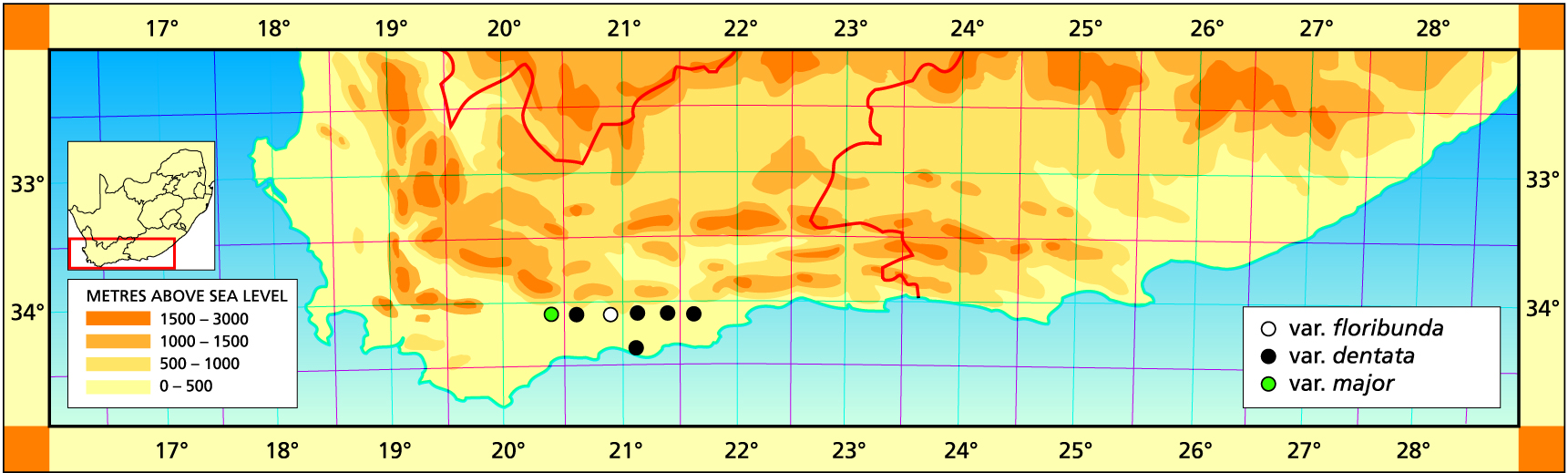
Distribution: 3420 (Bredasdorp): 6km N. Heidelberg (-BB), Smith 5545 (NBG, PRE), Bayer 158 (NBG); Heidelberg commonage (-BB), Ferguson in BOL20507.

Haworthia floribunda var. floribunda JDV88/22 north of Heidleberg. One of the few places where the plants have no spination. 
Haworthia floribunda var. floribunda JDV89/22 north of Heidelberg.
b. var. dentata var. nov.
Type: CAPE-3421 (Riversdale): W. Riversdale, Dekenah 90 in Smith 5502 (NBG, Holo.).
dentata: toothed.
Differs in being smaller, to 4cmφ, the leaves very dark green, slightly scabrid and with spined margins. (A var. floribunda foliis subtiliter scabridis et denti-marginatis differt).
This variety describes the smaller form which has distinct and widely spaced marginal spines. The leaves are very dark green and also slightly scabrid. It occurs from the Bontebok Park at Swellendam to northwest of Riversdale, and includes larger forms east of Riversdale also with pronounced marginal spines.
Distribution: 3420 (Bredasdorp): Bontebok Park (-BA), Bayer 3439 (NBG); E. Buffeljachts (-BA), Viviers 156 (NBG). 3421(Riversdale): W. Riversdale (-AA), Dekenah 90 in Smith 5502 (NBG): 5km N. Riversdale (-AB), Smith 5381 (NBG, PRE); 15km E. Riversdale (-AB), Smith 5758 (NBG); Dassieklip (-AC), Venter 92/31 (NBG); Wydersrivier (-BA), Smith 5491, 6781 (NBG), Bayer 2311.

Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Very difficult to find when dry. 
Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Showing the more scabrid leaves of this variety. 
Haworthia floribunda var. dentata JDV93/56 east of Riversdale. Also with the characteristic rounded leaf-tips. 
Haworthia floribunda var. dentata JDV94/47 north-east of Riversdale. Usually well-hidden and small. 
Haworthia floribunda var. dentata JDV93/58 east of Riversdale. A large robust darkest clone in cultivation. 
Haworthia floribunda var. dentata JDV93/58 east of Riversdale. In among stones and lichens.
c. var. viridescens var.nov.
Type: CAPE-3420 (Swellendam): S. Swellendam (-AB), De Kok (NBG, Holo.).
viridescens: becoming green.
Very green plants with darker coloration at the basal leaf margins. Relatively glabrous and more robust in cultivation. (A var. floribunda habitu robusto et colore viridi vivido ad margines basales foliorum atranti differt).
This is a large robust plant in cultivation and includes two forms, one of which has pointed leaves. The coloration is greener than normal for even cultivated plants of the species.
Distribution: 3420 (Bredasdorp): Below Swellendam Stn. (-AB)., C.A. Smith 2724a (PRE); S. Swellendam (-AB), D.De Kok (NBG).

Haworthia floribunda var. major JDV93/3 south of Swellendam. Plants in the field are quit small. 
Haworthia floribunda var. major JDV91/3 south of Swellendam. Leaves often without spines and tips sometimes pointed.







































































































































































































































































